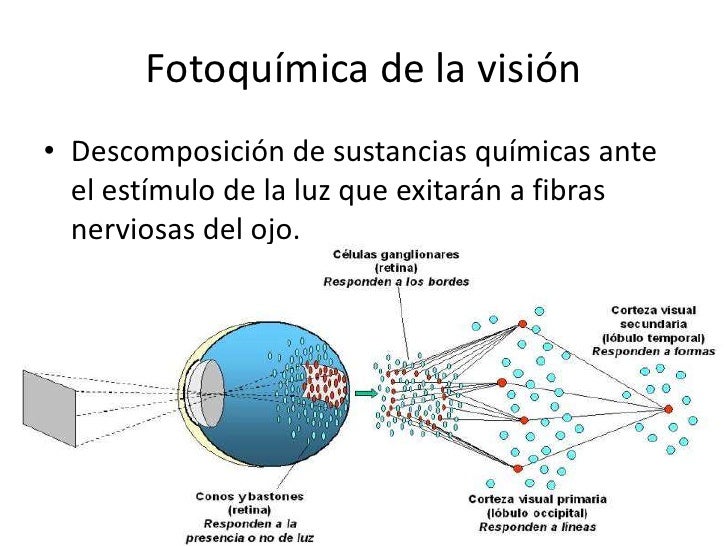
El corazón humano tiene un sistema especial para la autoexcitación rítmica y la contracción repetitiva aproximadamente 100.000 veces al día, o 3.000 millones de veces en una vida humana de duración media. Este impresionante logro es realizado por un sistema que: 1) genera impulsos eléctricos rítmicos para iniciar la contracción rítmica del músculo cardíaco 2) conduce estos estímulos rápidamente por todo el corazón. Cuando este sistema funciona normalmente, las aurículas se contraen aproximadamente 1/6 de segundo antes de la contracción ventricular, lo que permite el llenado de los ventrículos antes de que bombeen la sangre a través de los pulmones y de la circulación periférica. Este sistema rítmico y de conducción del corazón se puede lesionar en las cardiopatías, especialmente en la isquemia de los tejidos cardíacos que se debe a una disminución del flujo sanguíneo coronario. La consecuencia es con frecuencia una alteración del ritmo cardíaco o una secuencia anormal de contracción de las cavidades cardíacas, con una posible alteración grave de la eficacia de la función de bomba del corazón, incluso hasta el grado de producir la muerte.
Sistema de excitación especializado y de conducción del corazón. El nódulo sinusal (también denominado nódulo sinoauricular o SA), en el que se genera el impulso rítmico normal; las vías internodulares que conducen impulsos desde el nódulo sinusal hasta el nódulo auriculoventricular (AV); el nódulo AV, en el cual los impulsos originados en las aurículas se retrasan antes de penetrar en los ventrículos; el haz AV, que conduce impulsos desde las aurículas hacia los ventrículos, y las ramas izquierda y derecha del haz de fibras de Purkinje, que conducen los impulsos cardíacos por todo el tejido de los ventrículos.
Nódulo sinusal (sinoauricular) El nódulo sinusal (también denominado nódulo sinoauricular) es una banda elipsoide, aplanada y pequeña de músculo cardíaco especializado de aproximadamente 3 mm de anchura, 15 mm de longitud y 1 mm de grosor. Está localizado en la pared posterolateral superior de la aurícula derecha, inmediatamente inferior y ligeramente lateral a la desembocadura de la vena cava superior. Las fibras de este nódulo casi no tienen filamentos musculares contráctiles y cada una de ellas tiene solo de 3 a 5 μm de diámetro, en contraste con un diámetro de 10 a 15 μm para las fibras musculares auriculares circundantes, las fibras del nódulo sinusal se conectan directamente con las fibras musculares auriculares, de modo que todos los potenciales de acción que comienzan en el nódulo sinusal se propagan inmediatamente hacia la pared del músculo auricular.
Ritmicidad eléctrica automática de las fibras sinusales Algunas fibras cardíacas tienen la capacidad de autoexcitación, que es un proceso que puede producir descargas y contracciones rítmicas automáticas. Esta capacidad es especialmente cierta en el caso de las fibras del sistema especializado de conducción del corazón, entre ellas las fibras del nódulo sinusal. Por este motivo el nódulo sinusal habitualmente controla la frecuencia del latido de todo el corazón, como se analiza en detalle más adelante en este mismo capítulo. En primer lugar se va a describir esta ritmicidad automática. Mecanismo de la ritmicidad del nódulo sinusal Potenciales de acción registrados desde el interior de una fibra del nódulo sinusal durante tres latidos cardíacos y, a modo de comparación, un único potencial de acción de una fibra muscular ventricular, entre descargas el «potencial de membrana en reposo» de la fibra del nódulo sinusal tiene una negatividad de aproximadamente –55 a –60 mV, en comparación con –85 a –90 mV para la fibra muscular ventricular. La causa de esta menor negatividad es que las membranas celulares de las fibras sinusales son permeables naturalmente a los iones sodio y calcio, y las cargas positivas de los iones sodio y calcio que entran neutralizan parte de la negatividad intracelular.
El músculo cardíaco tiene tres tipos de canales iónicos de membrana que tienen funciones importantes en la generación de los cambios de voltaje en el potencial de acción. Los tipos son: 1) los canales rápidos de sodio. 2) los canales de calcio de tipo L (canales lentos de sodio-calcio). 3) los canales de potasio. La apertura de los canales rápidos de sodio durante algunas diezmilésimas de segundo es responsable de la rápida espiga ascendente del potencial de acción que se observa en el músculo ventricular, debido a la entrada rápida de iones sodio positivos hacia el interior de la fibra. La «meseta» del potencial de acción ventricular está producida principalmente por la apertura más lenta de los canales lentos de sodio-calcio, que dura aproximadamente 0,3 s. Finalmente, la apertura de los canales de potasio permite la difusión de grandes cantidades de iones potasio positivos hacia el exterior a través de la membrana de la fibra y devuelve el potencial de membrana a su nivel de reposo. Sin embargo, hay una diferencia en la función de estos canales en la fibra del nódulo sinusal porque el potencial «de reposo» es mucho menos negativo (de solo –55 mV en la fibra nodular, en lugar de los –90 mV de la fibra muscular ventricular). A este nivel de –55 mV, los canales rápidos de sodio principalmente ya se han «inactivado», lo que significa que han sido bloqueados. La causa de esto es que siempre que el potencial de membrana es menos negativo de aproximadamente –55 mV durante más de algunos milisegundos, las compuertas de inactivación del interior de la membrana celular que cierran los canales rápidos de sodio se cierran y permanecen de esta manera, solo se pueden abrir los canales lentos de sodio-calcio (es decir, se pueden «activar») y, por tanto, pueden producir el potencial de acción. En consecuencia, el potencial de acción del nódulo auricular se produce más lentamente que el potencial de acción del músculo ventricular. Además, después de la producción del potencial de acción, el regreso del potencial a su estado negativo también se produce lentamente, en lugar del regreso súbito que se produce en la fibra ventricular.
El nodo sinusal descarga con mayor rapidez que el Nodo AV o el Sistema His-Purkinje. Marcapasos anormales y ectópicos: Zonas de descarga rítmica mas rápida que la del nodo sinusal. El marcapaso se desplaza hacia esa zona (Nodo AV o Fibras de Purkinje). Un marcapaso en un lugar diferente del nodo sinusal es un Marcapaso Ectópico, hace que la contracción de las diferentes partes del cuerpo sea anormal. Bloqueo AV. Sindrome de Stokes-Adams.
Autoexcitación de las fibras del nódulo sinusal. Debido a la elevada concentración de iones sodio en el líquido extracelular en el exterior de la fibra nodular, así como al número moderado de canales de sodio abiertos previamente, los iones sodio positivos del exterior de las fibras normalmente tienden a desplazarse hacia el interior, entre los latidos cardíacos, la entrada de iones sodio de carga positiva produce una elevación lenta del potencial de membrana en reposo en dirección positiva, el potencial «en reposo» aumenta gradualmente y se hace menos negativo entre cada dos latidos sucesivos. Cuando el potencial alcanza un voltaje umbral de aproximadamente –40 mV, los canales de calcio de tipo L se «activan», produciendo de esta manera el potencial de acción. Por tanto, básicamente, la permeabilidad inherente de las fibras del nódulo sinusal a los iones sodio y calcio produce su autoexcitación. ¿Por qué esta permeabilidad a los iones sodio y calcio no hace que las fibras del nódulo sinusal permanezcan despolarizadas todo el tiempo? Durante el transcurso del potencial de acción se producen dos fenómenos que impiden dicho estado de despolarización constante. Primero, los canales de calcio de tipo L se inactivan (es decir, se cierran) en un plazo de aproximadamente 100 a 150 ms después de su apertura, y segundo, aproximadamente al mismo tiempo se abren números muy elevados de canales de potasio. Por tanto, se interrumpe el flujo de entrada de iones positivos calcio y sodio a través de los canales de calcio de tipo L, mientras que al mismo tiempo grandes cantidades de iones positivos de potasio difunden hacia el exterior de la fibra. Estos dos efectos reducen el potencial intracelular hasta devolverlo a su nivel de reposo negativo y, por tanto, ponen fin al potencial de acción. Además, los canales de potasio permanecen abiertos durante algunas décimas de segundo más, manteniendo transitoriamente el movimiento de cargas positivas hacia el exterior de la célula, con el consiguiente exceso de negatividad en el interior de la fibra; este proceso se denomina hiperpolarización. El estado de hiperpolarización inicialmente desplaza el potencial de membrana «en reposo» hacia abajo hasta aproximadamente –55 o –60 mV al final del potencial de acción. ¿Por qué este nuevo estado de hiperpolarización no se mantiene indefinidamente? El motivo es que en las décimas de segundo siguientes al final del potencial de acción se cierran cada vez más canales de potasio. Los iones sodio y calcio que fluyen hacia el interior una vez más compensan el flujo de salida de iones potasio, lo que lleva a que el potencial «de reposo» se desplace hacia arriba una vez más, alcanzando finalmente el nivel liminal de aproximadamente –40 mV. Después comienza de nuevo todo el proceso: autoexcitación para generar el potencial de acción, recuperación del potencial de acción, hiperpolarización después de que haya finalizado el potencial de acción, desplazamiento del potencial «de reposo» hasta el umbral y, finalmente, reexcitación para generar un nuevo ciclo. Este proceso continúa indefinidamente durante toda la vida de una persona. Las vías internodulares e interauriculares transmiten impulsos cardíacos a través de las aurículas Los extremos de las fibras del nódulo sinusal se conectan directamente con las fibras musculares auriculares circundantes. Por tanto, los potenciales de acción que se originan en el nódulo sinusal viajan hacia estas fibras musculares auriculares. De esta manera, el potencial de acción se propaga por toda la masa muscular auricular y, finalmente, llega hasta el nódulo AV. La velocidad de conducción en la mayor parte del músculo auricular es de aproximadamente 0,3 m/s, pero la conducción es más rápida, de aproximadamente 1 m/s, en varias pequeñas bandas de fibras auriculares. Una de estas bandas, denominada banda interauricular anterior, atraviesa las paredes anteriores de las aurículas para dirigirse hacia la aurícula izquierda. Además, otras tres bandas pequeñas se incurvan a través de las paredes auriculares anterior, lateral y posterior, y terminan en el nódulo AV; se denominan, respectivamente, vías internodulares anterior, media y posterior. La causa de la velocidad de conducción más rápida de estas bandas es la presencia de fibras de conducción especializadas. Estas fibras son similares a las «fibras de Purkinje» de los ventrículos, que conducen incluso más rápidamente y que se analizan del modo siguiente. El nódulo auriculoventricular retrasa la conducción del impulso desde las aurículas a los ventrículos El sistema de conducción auricular está organizado de modo que el impulso cardíaco no viaja desde las aurículas hacia los ventrículos demasiado rápidamente; este retraso da tiempo para que las aurículas vacíen su sangre hacia los ventrículos antes de que comience la contracción ventricular. El retraso de la transmisión hacia los ventrículos se produce principalmente en el nódulo AV y en sus fibras de conducción adyacentes. El nódulo AV está localizado en la pared posterolateral de la aurícula derecha, inmediatamente detrás de la válvula tricúspide, las diferentes partes de este nódulo, más sus conexiones con las fibras de las vías internodulares auriculares entrantes y el haz AV de salida. Los intervalos temporales aproximados en fracciones de segundo entre el comienzo inicial del impulso cardíaco en el nódulo sinusal y su posterior aparición en el sistema del nódulo AV. Obsérvese que el impulso, después de viajar por las vías internodulares, llega al nódulo AV aproximadamente 0,03 s después de su origen en el nódulo sinusal. Después hay un retraso de otros 0,09 s en el propio nódulo AV antes de que el impulso entre en la porción penetrante del haz AV, a través del cual pasa hacia los ventrículos. Se produce un retraso final de otros 0,04 s principalmente en este haz AV penetrante, que está formado por múltiples fascículos pequeños que atraviesan el tejido fibroso que separa las aurículas de los ventrículos.
Sistema de conducción. Nodo SA. 0 seg. Inicio. Vías internodales seg. De retraso. Nodo AV. 0.9 seg. de retraso. Haz de His. 0.4 seg. de retraso seg de retraso, Así, el retraso total en el nódulo AV y en el sistema de AV es de aproximadamente 0,13 s. Este retraso, añadido al retraso inicial de la conducción de 0,03 s desde el nódulo sinusal hasta el nódulo AV, hace que haya un retraso total de 0,16 s antes de que la señal excitadora llegue finalmente al músculo ventricular que se está contrayendo.
Causa de la conducción lenta La conducción lenta en las fibras transicionales, nodulares y penetrantes del haz AV está producida principalmente por la disminución del número de uniones en hendidura entre células sucesivas de las vías de conducción, de modo que hay una gran resistencia a la conducción de los iones excitadores desde una fibra de conducción hasta la siguiente. Por tanto, es fácil ver por qué se tarda en excitar células sucesivas. Transmisión rápida en el sistema de Purkinje ventricular Las fibras de Purkinje especiales se dirigen desde el nódulo AV a través del haz AV hacia los ventrículos. Excepto en la porción inicial de estas fibras, donde penetran en la barrera fibrosa AV, tienen características funcionales bastante distintas a las de las fibras del nódulo AV. Son fibras muy grandes, incluso mayores que las fibras musculares ventriculares normales, y transmiten potenciales de acción a una velocidad de 1,5 a 4 m/s, una velocidad aproximadamente seis veces mayor que la del músculo ventricular normal y 150 veces mayor que la de algunas de las fibras del nódulo AV. Esta velocidad permite una transmisión casi instantánea del impulso cardíaco por todo el resto del músculo ventricular. Se piensa que la rápida transmisión de los potenciales de acción por las fibras de Purkinje está producida por un gran aumento del nivel de permeabilidad de las uniones en hendidura de los discos intercalados entre las células sucesivas que componen las fibras de Purkinje. Por tanto, los iones pasan fácilmente de una célula a la siguiente, aumentando de esta manera la velocidad de la transmisión. Las fibras de Purkinje también tienen muy pocas miofibrillas, lo que significa que se contraen poco o nada durante la transmisión de los impulsos. Conducción unidireccional a través del haz AV Una característica especial del haz AV es la imposibilidad, excepto en estados anormales, de que los potenciales de acción viajen retrógradamente desde los ventrículos hacia las aurículas. Esta característica impide la reentrada de los impulsos cardíacos por esta ruta desde los ventrículos hacia las aurículas, permitiendo solo la contracción anterógrada desde las aurículas hacia los ventrículos. Además, se debe recordar que en todas las localizaciones excepto en el haz AV el músculo auricular está separado del músculo ventricular por una barrera fibrosa continua, de la que se muestra una porción. Esta barrera normalmente actúa como aislante para impedir el paso de los impulsos cardíacos entre el músculo auricular y ventricular a través de cualquier ruta distinta a la conducción anterógrada a través del propio haz AV. (En casos infrecuentes un puente muscular anormal penetra en la barrera fibrosa en otra localización distinta al haz AV. En estas condiciones el impulso cardíaco puede entrar en las aurículas desde los ventrículos y producir arritmias cardíacas graves.) Distribución de las fibras de Purkinje en los ventrículos: las ramas izquierda y derecha del haz Después de penetrar en el tejido fibroso que está entre el músculo auricular y ventricular, la porción distal del haz AV se dirigía hacia abajo en el interior del tabique interventricular a lo largo de 5 a 15 mm hacia la punta del corazón, después el haz se divide en las ramas izquierda y derecha del haz, que están debajo del endocardio en los dos lados respectivos del tabique interventricular. Cada una de las ramas se dirige hacia abajo, hacia la punta del ventrículo, dividiéndose progresivamente en ramas más pequeñas. Estas ramas, a su vez, siguen un trayecto en dirección lateral alrededor de cada una de las cavidades ventriculares y hacia atrás, hacia la base del corazón. Los extremos de las fibras de Purkinje penetran en aproximadamente un tercio del grosor de la masa muscular y finalmente se continúan con las fibras musculares cardíacas. Desde el momento en el que el impulso cardíaco entre las ramas del haz en el tabique interventricular hasta que sale de las terminaciones de las fibras de Purkinje el tiempo total transcurrido es en promedio de solo 0,03 s. Por tanto, una vez que el impulso cardíaco ha entrado en el sistema de conducción ventricular de Purkinje, se propaga casi inmediatamente a toda la masa del músculo ventricular. Transmisión del impulso cardíaco en el músculo ventricular. Una vez que el impulso llega a los extremos de las fibras de Purkinje se transmite a través de la masa del músculo ventricular por las propias fibras musculares ventriculares. La velocidad de transmisión es ahora solo de 0,3 a 0,5 m/s, una sexta parte de la velocidad de las fibras de Purkinje. El músculo cardíaco envuelve el corazón en una doble espiral, con tabiques fibrosos entre las capas en espiral; por tanto, el impulso cardíaco no viaja necesariamente directamente hacia el exterior, hacia la superficie del corazón, sino que se angula hacia la superficie a lo largo de las direcciones de las espirales. Debido a esta inclinación, la transmisión desde la superficie endocárdica a la superficie epicárdica del ventrículo precisa hasta otros 0,03 s, aproximadamente igual al tiempo necesario para la transmisión por toda la porción ventricular del sistema de Purkinje. Así, el tiempo total para la transmisión del impulso cardíaco desde las ramas iniciales del haz hasta las últimas fibras del músculo ventricular en el corazón normal es de aproximadamente 0,06 s. Resumen de la propagación del impulso cardíaco a través del corazón La transmisión del impulso cardíaco en el corazón humano. Los números de la figura representan los intervalos de tiempo, en fracciones de segundo, que transcurren desde el origen del impulso cardíaco en el nódulo sinusal hasta su aparición en cada uno de los puntos respectivos del corazón. Obsérvese que el impulso se propaga a una velocidad moderada a través de las aurículas, aunque se retrasa más de 0,1 s en la región del nódulo AV antes de aparecer en el haz AV del tabique interventricular. Una vez que ha entrado en este haz, se propaga muy rápidamente a través de las fibras de Purkinje por todas las superficies endocárdicas de los ventrículos. Después el impulso se propaga de nuevo algo más lentamente a través del músculo ventricular hacia las superficies epicárdicas.
Control de la excitación y la conducción en el corazón El nódulo sinusal es el marcapasos normal del corazón El impulso normalmente se origina en el nódulo sinusal. En algunas situaciones anormales no ocurre así. Otras partes del corazón pueden presentar también una excitación rítmica intrínseca de la misma forma que lo hacen las fibras del nódulo sinusal; esta capacidad es particularmente cierto en el caso de las fibras del nódulo AV y de las fibras de Purkinje. Las fibras del nódulo AV, cuando no son estimuladas por alguna fuente externa, descargan a una frecuencia rítmica intrínseca de 40 a 60 veces por minuto, y las fibras de Purkinje lo hacen a una frecuencia de entre 15 y 40 veces por minuto. Estas frecuencias son distintas a la frecuencia normal del nódulo sinusal, de 70 a 80 veces por minuto. ¿Por qué entonces es el nódulo sinusal, y no el nódulo AV ni las fibras de Purkinje, el que controla la ritmicidad del corazón? La respuesta procede del hecho de que la frecuencia de descarga del nódulo sinusal es considerablemente mayor que la frecuencia de descarga autoexcitadora natural de las fibras del nódulo AV y de las fibras de Purkinje. Cada vez que se produce una descarga en el nódulo sinusal su impulso se conduce hacia el nódulo AV y hacia las fibras de Purkinje, produciendo también la descarga de sus membranas. Sin embargo, el nódulo sinusal produce una nueva descarga antes de que las fibras del nódulo AV o las fibras de Purkinje puedan alcanzar sus propios umbrales de autoexcitación. Por tanto, el nuevo impulso procedente del nódulo sinusal descarga tanto las fibras del nódulo AV como las fibras de Purkinje antes de que se pueda producir autoexcitación en cualquiera de esas estructuras. Así, el nódulo sinusal controla el latido del corazón porque su frecuencia de descarga rítmica es más rápida que la de cualquier otra parte del corazón. Por tanto, el nódulo sinusal es casi siempre el marcapasos del corazón normal. Marcapasos anormales: marcapasos «ectópico» De manera ocasional alguna otra parte del corazón muestra una frecuencia de descarga rítmica que es más rápida que la del nódulo sinusal. Por ejemplo, a veces se produce este desarrollo en el nódulo AV o en las fibras de Purkinje cuando una de estas estructuras se altera. En ambos casos el marcapasos del corazón se desplaza desde el nódulo sinusal hasta el nódulo AV o las fibras de Purkinje excitadas. En casos menos frecuentes todavía, algún punto del músculo auricular o ventricular presenta una excitabilidad excesiva y se convierte en el marcapasos. Un marcapasos que está situado en una localización distinta al nódulo sinusal se denomina marcapasos «ectópico». Un marcapasos ectópico da lugar a una secuencia anormal de contracción de las diferentes partes del corazón y puede producir una debilidad significativa del bombeo cardíaco. Otra causa de desplazamiento del marcapasos es el bloqueo de la transmisión del impulso cardíaco desde el nódulo sinusal a las demás partes del corazón. El nuevo marcapasos se produce en este caso con más frecuencia en el nódulo AV o en la porción penetrante del haz AV en su trayecto hacia los ventrículos. Cuando se produce un bloqueo AV, es decir, cuando el impulso cardíaco no puede pasar desde las aurículas hacia los ventrículos a través del sistema del nódulo AV y del haz, las aurículas siguen latiendo a la frecuencia normal del ritmo del nódulo sinusal, mientras que habitualmente aparece un nuevo marcapasos en el sistema de Purkinje de los ventrículos que activa el músculo ventricular a una frecuencia de entre 15 y 40 latidos/min. Después de un bloqueo súbito del haz AV el sistema de Purkinje no comienza a emitir sus impulsos rítmicos intrínsecos hasta 5 a 20 s después porque, antes del bloqueo, las fibras de Purkinje habían estado «sobreexcitadas» por los rápidos impulsos sinusales y, en consecuencia, están en un estado suprimido. Durante estos 5 a 20 s los ventrículos dejan de bombear sangre y la persona se desvanece después de los primeros 4 a 5 s debido a la ausencia de flujo sanguíneo cerebral. Este retraso de la recuperación del corazón se denomina síndrome de StokesAdams. Si el período de retraso es demasiado largo, se puede producir la muerte. Importancia del sistema de Purkinje en la generación de una contracción sincrónica del músculo ventricular La rápida conducción del sistema de Purkinje permite normalmente que el impulso cardíaco llegue a casi todas las porciones de los ventrículos en un breve intervalo de tiempo, excitando la primera fibra muscular ventricular solo 0,03 a 0,06 s antes de la excitación de la última. Esta sincronización hace que todas las porciones del músculo de los dos ventrículos comiencen a contraerse casi al mismo tiempo y que después sigan contrayéndose durante aproximadamente otros 0,3 s. La función de bomba eficaz de las dos cavidades ventriculares precisa este tipo sincrónico de contracción. Si el impulso cardíaco viajara lentamente a través de los ventrículos, buena parte de la masa ventricular se contraería antes de la contracción del resto, en cuyo caso se produciría una gran disminución de la función global de bomba. De hecho, en algunos tipos de trastornos cardíacos, algunos de los cuales se analizan en los capítulos 12 y 13, se produce una transmisión lenta, y la eficacia del bombeo de los ventrículos disminuye hasta el 20-30%. Los nervios simpáticos y parasimpáticos controlan el ritmo cardíaco y la conducción de impulsos por los nervios cardíacos El corazón está inervado por nervios simpáticos y parasimpáticos. Los nervios parasimpáticos (vagos) se distribuyen principalmente a los nódulos SA y AV, en mucho menor grado al músculo de las dos aurículas y apenas directamente al músculo ventricular. Por el contrario, los nervios simpáticos se distribuyen en todas las regiones del corazón, con una intensa representación en el músculo ventricular, así como en todas las demás zonas. La estimulación parasimpática (vagal) ralentiza el ritmo y la conducción cardíacos La estimulación de los nervios parasimpáticos que llegan al corazón (los vagos) hace que se libere la hormona acetilcolina en las terminaciones nerviosas. Esta hormona tiene dos efectos principales sobre el corazón. Primero, reduce la frecuencia del ritmo del nódulo sinusal, y segundo, reduce la excitabilidad de las fibras de la unión AV entre la musculatura auricular y el nódulo AV, retrasando de esta manera la transmisión del impulso cardíaco hacia los ventrículos. Una estimulación vagal débil a moderada reduce la frecuencia del bombeo del corazón, con frecuencia hasta un valor tan bajo como la mitad de lo normal. Además, la estimulación intensa de los nervios vagos puede interrumpir completamente la excitación rítmica del nódulo sinusal o puede bloquear completamente la transmisión del impulso cardíaco desde las aurículas hacia los ventrículos a través del nódulo AV. En cualquiera de los casos, las señales excitadores rítmicas ya no se transmiten hacia los ventrículos. Los ventrículos pueden dejar de latir durante 5 a 20 s, pero después alguna área pequeña de las fibras de Purkinje, habitualmente en la porción del tabique interventricular del haz AV, presenta un ritmo propio y genera la contracción ventricular a una frecuencia de 15 a 40 latidos/min. Este fenómeno se denomina escape ventricular. Mecanismo de los efectos vagales La acetilcolina que se libera en las terminaciones nerviosas vagales aumenta mucho la permeabilidad de las membranas de las fibras a los iones potasio, lo que permite la salida rápida de potasio desde las fibras del sistema de conducción. Este proceso da lugar a un aumento de la negatividad en el interior de las fibras, un efecto que se denomina hiperpolarización, que hace que este tejido excitable sea mucho menos excitable. En el nódulo sinusal, el estado de hiperpolarización hace el potencial de membrana «en reposo» de las fibras del nódulo sinusal mucho más negativo de lo habitual, es decir, de –65 a –75 mV en lugar del nivel normal de –55 a –60 mV. Por tanto, el aumento inicial del potencial de membrana del nódulo sinusal que produce la corriente de entrada de sodio y de calcio tarda mucho más en alcanzar el potencial liminal para la excitación. Este requisito retrasa mucho la frecuencia de ritmicidad de estas fibras nodulares. Si la estimulación vagal es lo suficientemente intensa es posible detener totalmente la autoexcitación rítmica de este nódulo. En el nódulo AV, el estado de hiperpolarización producido por la estimulación vagal hace que sea difícil que las pequeñas fibras auriculares que entran en el nódulo generen una corriente de una intensidad suficiente como para excitar las fibras nodulares. Por tanto, el factor de seguridad para la transmisión del impulso cardíaco a través de las fibras de transición hacia las fibras del nódulo AV disminuye. Una reducción moderada simplemente retrasa la conducción del impulso, aunque una disminución grande bloquea totalmente la conducción. La estimulación simpática aumenta el ritmo y la conducción del corazón La estimulación simpática produce esencialmente los efectos contrarios sobre el corazón a los que produce la estimulación vagal, como se señala a continuación. Primero, aumenta la frecuencia de descarga del nódulo sinusal. Segundo, aumenta la velocidad de conducción, así como el nivel de excitabilidad de todas las porciones del corazón. Tercero, aumenta mucho la fuerza de contracción de toda la musculatura cardíaca, tanto auricular como ventricular. En breve, la estimulación simpática aumenta la actividad global del corazón. La estimulación máxima casi puede triplicar la frecuencia del latido cardíaco y puede aumentar la fuerza de la contracción del corazón hasta dos veces. Mecanismo del efecto simpático La estimulación de los nervios simpáticos libera la hormona noradrenalina en las terminaciones nerviosas simpáticas. La noradrenalina estimula, a su vez, los receptores β1-adrenérgicos, que median en los efectos sobre la frecuencia cardíaca. No está del todo claro el mecanismo preciso mediante el que la estimulación β1-adrenérgica actúa sobre las fibras del músculo cardíaco, aunque se piensa que aumenta la permeabilidad de la membrana de las fibras a los iones sodio y calcio. En el nódulo sinusal, un aumento de la permeabilidad a sodio-calcio genera un potencial en reposo más positivo y también produce un aumento de la velocidad del ascenso del potencial de membrana diastólico hacia el nivel liminal para la autoexcitación, acelerando de esta forma la autoexcitación y, por tanto, aumentando la frecuencia cardíaca. En el nódulo AV y en los haces AV, el aumento de la permeabilidad a sodio-calcio hace que sea más fácil que el potencial de acción excite todas las porciones sucesivas de los haces de las fibras de conducción, disminuyendo de esta manera el tiempo de conducción desde las aurículas hasta los ventrículos. El aumento de la permeabilidad a los iones calcio es responsable al menos en parte del aumento de la fuerza contráctil del músculo cardíaco bajo la influencia de la estimulación simpática, porque los iones calcio tienen una función importante en la excitación del proceso contráctil de las miofibrillas.
El corazón está formado realmente por dos bombas separadas: un corazón derecho que bombea sangre hacia los pulmones y un corazón izquierdo que bombea sangre a través de la circulación sistémica que aporta flujo sanguíneo a los demás órganos y tejidos del cuerpo. A su vez, cada uno de estos corazones es una bomba bicameral pulsátil formada por una aurícula y un ventrículo. Cada una de las aurículas es una bomba débil de cebado del ventrículo, que contribuye a transportar sangre hacia el ventrículo correspondiente. Los ventrículos después aportan la principal fuerza del bombeo que impulsa la sangre: 1) hacia la circulación pulmonar por el ventrículo derecho, o 2) hacia la circulación sistémica por el ventrículo izquierdo..
«FISIOLOGIA DEL MUSCULO CARDIACO» El corazón está formado por tres tipos principales de músculo cardíaco: músculo auricular, músculo ventricular y fibras musculares especializadas de excitación y de conducción. El músculo auricular y ventricular se contrae de manera muy similar al músculo esquelético, excepto porque la duración de la contracción es mucho mayor. No obstante, las fibras especializadas de excitación y de conducción del corazón se contraen solo débilmente porque contienen pocas fibrillas contráctiles; en cambio, presentan descargas eléctricas rítmicas automáticas en forma de potenciales de acción o conducción de los potenciales de acción por todo el corazón, formando así un sistema excitador que controla el latido rítmico cardíaco. «ANATOMÍA FISIOLÓGICA DEL MÚSCULO CARDÍACO.» Las zonas oscuras que atraviesan las fibras musculares cardíacas se denominan discos intercalados; realmente son membranas celulares que separan las células musculares cardíacas individuales entre sí. Es decir, las fibras musculares cardíacas están formadas por muchas células individuales conectadas entre sí en serie y en paralelo. En cada uno de los discos intercalados las membranas celulares se fusionan entre sí para formar uniones «comunicantes» (en hendidura) permeables que permiten una rápida difusión. Por tanto, desde un punto de vista funcional los iones se mueven con facilidad en el líquido intracelular a lo largo del eje longitudinal de las fibras musculares cardíacas, de modo que los potenciales de acción viajan fácilmente desde una célula muscular cardíaca a la siguiente, a través de los discos intercalados. Por tanto, el músculo cardíaco es un sincitio de muchas células musculares cardíacas en el que las células están tan interconectadas entre sí que cuando una célula se excita el potencial de acción se propaga rápidamente a todas. El corazón realmente está formado por dos sincitios: el sincitio auricular, que forma las paredes de las dos aurículas, y el sincitio ventricular, que forma las paredes de los dos ventrículos. Las aurículas están separadas de los ventrículos por tejido fibroso que rodea las aberturas de las válvulas auriculoventriculares (AV) entre las aurículas y los ventrículos. Normalmente los potenciales no se conducen desde el sincitio auricular hacia el sincitio ventricular directamente a través de este tejido fibroso. Por el contrario, solo son conducidos por medio de un sistema de conducción especializado denominado haz AV, que es un fascículo de fibras de conducción de varios milímetros de diámetro.
Esta división del músculo del corazón en dos sincitios funcionales permite que las aurículas se contraigan un pequeño intervalo antes de la contracción ventricular, lo que es importante para la eficacia del bombeo del corazón.
POTENCIALES DE ACCIÓN EN EL MÚSCULO CARDÍACO. El potencial de acción que se origina en la fibra ventricular es en promedio de aproximadamente 105 mV, lo que significa que el potencial intracelular aumenta desde un valor muy negativo, de aproximadamente –85 mV, entre los latidos hasta un valor ligeramente positivo, de aproximadamente +20 mV, durante cada latido. Después de la espiga inicial la membrana permanece despolarizada durante aproximadamente 0,2 s, mostrando una meseta, seguida al final de la meseta de una repolarización súbita. La presencia de esta meseta del potencial de acción ha- ce que la contracción ventricular dure hasta 15 veces más en el músculo cardíaco que en el músculo esquelético. En el músculo cardíaco, el potencial de acción está producido por la apertura de dos tipos de canales: 1) los mismos canales rápidos de sodio activados por el voltaje que en el músculo esquelético 2) otra población totalmente distinta de canales de calcio de tipo L (canales lentos de calcio), que también se denominan canales de calcio-sodio. Esta segunda población de canales difiere de los canales rápidos de sodio en que se abren con mayor lentitud y, lo que es incluso más importante, permanecen abiertos durante varias décimas de segundo. Durante este tiempo fluye una gran cantidad de iones tanto calcio como sodio a través de estos canales hacia el interior de la fibra muscular cardíaca, y esta actividad mantiene un período prolongado de despolarización, dando lugar a la meseta del potencial de acción. Además, los iones calcio que entran durante esta fase de meseta activan el proceso contráctil del músculo, mientras que los iones calcio que producen la contracción del músculo esquelético proceden del retículo sarcoplásmico intracelular. La segunda diferencia funcional importante entre el músculo cardíaco y el músculo esquelético que ayuda a explicar tanto el potencial de acción prolongado como su meseta es la siguiente: inmediatamente después del inicio del potencial de acción, la permeabilidad de la membrana del músculo cardíaco a los iones potasio disminuye aproximadamente cinco veces, un efecto que no aparece en el músculo esquelético. Esta disminución de la permeabilidad al potasio se puede deber al exceso de flujo de entrada de calcio a través de los canales de calcio que se acaba de señalar. Independientemente de la causa, la disminución de la permeabilidad al potasio reduce mucho el flujo de salida de iones potasio de carga positiva durante la meseta del potencial de acción y, por tanto, impide el regreso rápido del voltaje del potencial de acción a su nivel de reposo. Cuando los canales lentos de calcio-sodio se cierran después de 0,2 a 0,3 s y se interrumpe el flujo de entrada de iones calcio y sodio, también aumenta rápidamente la permeabilidad de la membrana a los iones potasio; esta rápida pérdida de potasio desde la fibra inmediatamente devuelve el potencial de membrana a su nivel de reposo, finalizando de esta manera el potencial de acción.
Potencial de acción en una fibra de Purkinje
«FASES DEL POTENCIAL DE ACCION DEL MUSCULO CARDIACO»
Fase 0 (despolarización) –> los canales de sodio rápidos se abren. Cuando la célula cardíaca es estimulada y se despolariza, el potencial de membrana se hace más positivo. Los canales de sodio activados por el voltaje (canales de sodio rápidos) se abren y permiten que el sodio circule rápidamente hacia el interior de la célula y la despolarice. El potencial de membrana alcanza +20 mV aproximadamente antes de que los canales de sodio se cierren. Fase 1 (repolarización inicial) –> los canales de sodio rápidos se cierran. Los canales de sodio se cierran, la célula empieza a repolarizarse y los iones potasio salen de la célula a través de los canales de potasio. Fase 2 (meseta) –> los canales de calcio se abren y los canales de potasio rápidos se cierran. Tiene lugar una breve repolarización inicial y el potencial de acción alcanza una meseta como consecuencia de: 1) una mayor permeabilidad a los iones calcio 2) la disminución de la permeabilidad a los iones potasio. Los canales de calcio activados por el voltaje se abren lentamente durante las fases 1 y 0, y el calcio entra en la célula. Después, los canales de potasio se cierran, y la combinación de una reducción en la salida de iones potasio y un aumento de la entrada de iones calcio lleva a que el potencial de acción alcance una meseta. Fase 3 (repolarización rápida) –> los canales de calcio se cierran y los canales de potasio lentos se abren. El cierre de los canales iónicos de calcio y el aumento de la permeabilidad a los iones potasio, que permiten que los iones potasio salgan rápidamente de la célula, pone fin a la meseta y devuelve el potencial de membrana de la célula a su nivel de reposo. Fase 4 (potencial de membrana de reposo) –> con valor medio aproximado de −90 mV. La velocidad de la conducción de la señal del potencial de acción excitador a lo largo de las fibras musculares auriculares y ventriculares es de unos 0,3 a 0,5 m/s, o aproximadamente 1/250 de la velocidad en las fibras nerviosas grandes y en torno a 1/10 de la velocidad en las fibras musculares esqueléticas. El período refractario normal del ventrículo es de 0,25 a 0,30 s, que es aproximadamente la duración del potencial de acción en meseta prolongado. Hay un período refractario relativo adicional de aproximadamente 0,05 s, durante el cual es más difícil de lo normal excitar el músculo pero, sin embargo, se puede excitar con una señal excitadora muy intensa, como se demuestra por la contracción «prematura» temprana del segundo.
Acoplamiento excitación-contracción: función de los iones calcio y de los túbulos transversos El término «acoplamiento excitación-contracción» se refiere al mecanismo mediante el cual el potencial de acción hace que las miofibrillas del músculo se contraigan, cuando un potencial de acción pasa sobre la membrana del músculo cardíaco el potencial de acción se propaga hacia el interior de la fibra muscular cardíaca a lo largo de las membranas de los túbulos transversos (T). Los potenciales de acción de los túbulos T, a su vez, actúan sobre las membranas de los túbulos sarcoplásmicos longitudinales para producir la liberación de iones calcio hacia el sarcoplasma muscular desde el retículo sarcoplásmico. En algunas milésimas de segundo más estos iones calcio difunden hacia las miofibrillas y catalizan las reacciones químicas que favorecen el deslizamiento de los filamentos de actina y de miosina entre sí, lo que da lugar a la contracción muscular.
Sin el calcio procedente de los túbulos T la fuerza de la contracción del músculo cardíaco se reduciría de manera considerable porque el retículo sarcoplásmico del músculo cardíaco está peor desarrollado que el del músculo esquelético y no almacena suficiente calcio para generar una contracción completa. No obstante, los túbulos T del músculo cardíaco tienen un diámetro cinco veces mayor que los túbulos del músculo esquelético, lo que significa un volumen 25 veces mayor. Además, en el interior de los túbulos T hay una gran cantidad de mucopolisacáridos que tienen carga negativa y que se unen a una abundante reserva de iones calcio, manteniéndolos disponibles para su difusión hacia el interior de la fibra muscular cardíaca cuando aparece un potencial de acción en un túbulo T. La fuerza de la contracción del músculo cardíaco depende en gran medida de la concentración de iones calcio en los líquidos extracelulares. De hecho, un corazón situado en una solución sin calcio dejará rápidamente de latir. La razón de esta respuesta es que las aberturas de los túbulos T atraviesan directamente la membrana de la célula muscular cardíaca hacia los espacios extracelulares que rodean las células, lo que permite que el mismo líquido extracelular que está en el intersticio del músculo cardíaco se introduzca en los túbulos T. En consecuencia, la cantidad de iones calcio en el sistema de los túbulos T (es decir, la disponibilidad de iones calcio para producir la contracción del músculo cardíaco) depende en gran medida de la concentración de iones calcio en el líquido extracelular. En cambio, la fuerza de la contracción del músculo esquelético apenas se ve afectada por cambios moderados de la concentración de calcio en el líquido extracelular porque la contracción del músculo esquelético está producida casi por completo por los iones calcio que son liberados por el retículo sarcoplásmico del interior de la propia fibra muscular esquelética. Al final de la meseta del potencial de acción cardíaco se interrumpe súbitamente el flujo de entrada de iones calcio hacia el interior de la fibra muscular y los iones calcio del sarcoplasma se bombean rápidamente hacia el exterior de las fibras musculares, hacia el retículo sarcoplásmico y hacia el espacio de los túbulos T-líquido extracelular. El transporte de calcio de nuevo al retículo sarcoplásmico se consigue con la ayuda de una bomba de calcio adenosina trifosfatasa (ATPasa). Los iones calcio se eliminan también de la célula mediante un intercambiador de sodiocalcio. El sodio que entra en la célula durante este intercambio se transporta después fuera de la célula por acción de la bomba de sodio-potasio ATPasa. En consecuencia, se interrumpe la contracción hasta que llega un nuevo potencial de acción.
El músculo cardíaco comienza a contraerse algunos milisegundos después de la llegada del potencial de acción y sigue contrayéndose hasta algunos milisegundos después de que finalice. Por tanto, la duración de la contracción del músculo cardíaco depende principalmente de la duración del potencial de acción, incluyendo la meseta, aproximadamente 0,2 s en el músculo auricular y 0,3 s en el músculo ventricular.
«CICLO CARDIACO»
Los fenómenos cardíacos que se producen desde el comienzo de un latido cardíaco hasta el comienzo del siguiente se denominan ciclo cardíaco. Cada ciclo es iniciado por la generación espontánea de un potencial de acción en el nódulo sinusal. Este nódulo está localizado en la pared superolateral de la aurícula derecha, cerca del orificio de la vena cava superior, y el potencial de acción viaja desde aquí rápidamente por ambas aurículas y después a través del haz AV hacia los ventrículos. Debido a esta disposición especial del sistema de conducción desde las aurículas hacia los ventrículos, hay un retraso de más de 0,1 s durante el paso delimpulso cardíaco desde las aurículas a los ventrículos. Esto permite que las aurículas se contraigan antes de la contracción ventricular, bombeando de esta manera sangre hacia los ventrículos antes de que comience la intensa contracción ventricular. Por tanto, las aurículas actúan como bombas de cebado para los ventrículos, y los ventrículos a su vez proporcionan la principal fuente de potencia para mover la sangre a través del sistema vascular del cuerpo.
DIASTOLE Y SISTOLE. El ciclo cardíaco está formado por un período de relajación que se denomina diástole, seguido de un período de contracción denominado sístole. La duración del ciclo cardíaco total, incluidas la sístole y la diástole, es el valor inverso de la frecuencia cardíaca. Por ejemplo, si la frecuencia cardíaca es de 72 latidos/min, la duración del ciclo cardíaco es de 1/72 min/latido, aproximadamente 0,0139 min por latido, o 0,833 s por latido.
Cuando aumenta la frecuencia cardíaca, la duración de cada ciclo cardíaco disminuye, incluidas las fases de contracción y relajación. La duración del potencial de acción y el período de contracción (sístole) también decrece, aunque no en un porcentaje tan elevado como en la fase de relajación (diástole). Para una frecuencia cardíaca normal de 72 latidos/min, la sístole comprende aproximadamente 0,4 del ciclo cardíaco completo. Para una frecuencia cardíaca triple de lo normal, la sístole supone aproximadamente 0,65 del ciclo cardíaco completo. Esto significa que el corazón que late a una frecuencia muy rápida no permanece relajado el tiempo suficiente para permitir un llenado completo de las cámaras cardíacas antes de la siguiente contracción.
Relación del electrocardiograma con el ciclo cardíaco
El electrocardiograma de la figura 9-7 muestra las ondas P, Q, R, S y T, que se analizan en los capítulos 11, 12 y 13. Son los voltajes eléctricos que genera el corazón, y son registrados mediante el electrocardiógrafo desde la superficie del cuerpo. La onda P está producida por la propagación de la despolarización en las aurículas, y es seguida por la contracción auricular, que produce una ligera elevación de la curva de presión auricular inmediatamente después de la onda P electrocardiográfica. Aproximadamente 0,16 s después del inicio de la onda P, las ondas QRS aparecen como consecuencia de la despolarización eléctrica de los ventrículos, que inicia la contracción de los ventrículos y hace que comience a elevarse la presión ventricular. Por tanto, el complejo QRS comienza un poco antes del inicio de la sístole ventricular. Finalmente, la onda T ventricular representa la fase de repolarización de los ventrículos, cuando las fibras del músculo ventricular comienzan a relajarse. Por tanto, la onda T se produce un poco antes del final de la contracción ventricular.
La sangre normalmente fluye de forma continua desde las grandes ventas hacia las aurículas; aproximadamente el 80% de la sangre fluye directamente a través de las aurículas hacia los ventrículos incluso antes de que se contraigan las aurículas. Después, la contracción auricular habitualmente produce un llenado de un 20% adicional de los ventrículos. La onda a está producida por la contracción auricular. Habitualmente la presión auricular derecha aumenta de 4 a 6 mmHg durante la contracción auricular y la presión auricular izquierda aumenta aproximadamente de 7 a 8 mmHg. La onda c se produce cuando los ventrículos comienzan a contraerse; está producida en parte por un ligero flujo retrógrado de sangre hacia las aurículas al comienzo de la contracción ventricular, pero principalmente por la protrusión de las válvulas AV retrógradamente hacia las aurículas debido al aumento de presión de los ventrículos. La onda v se produce hacia el final de la contracción ventricular; se debe al flujo lento de sangre hacia las aurículas desde las venas mientras las válvulas AV están cerradas durante la contracción ventricular. Después, cuando ya ha finalizado la contracción ventricular, las válvulas AV se abren, y permiten que esta sangre auricular almacenada fluya rápidamente hacia los ventrículos, lo que hace que la onda v desaparezca.
VENTRICULOS COMO BOMBAS.
Durante la sístole ventricular se acumulan grandes cantidades de sangre en las aurículas derecha e izquierda porque las válvulas AV están cerradas. Por tanto, tan pronto como ha finalizado la sístole y las presiones ventriculares disminuyen de nuevo a sus valores diastólicos bajos, el aumento moderado de presión que se ha generado en las aurículas durante la sístole ventricular inmediatamente abre las válvulas AV y permite que la sangre fluya rápidamente hacia los ventrículos, como se muestra en la elevación de la curva de volumen ventricular izquierdo, es el denominado período de llenado rápido de los ventrículos.
Desbordamiento de los ventrículos durante la sístole Período de contracción isovolumétrica (isométrica) Inmediatamente después del comienzo de la contracción ventricular se produce un aumento súbito de presión ventricular, lo que hace que se cierren las válvulas AV. Después son necesarios otros 0,02 a 0,03 s para que el ventrículo acumule una presión suficiente para abrir las válvulas AV semilunares (aórtica y pulmonar) contra las presiones de la aorta y de la arteria pulmonar. Por tanto, durante este período se produce contracción en los ventrículos, pero no se produce vaciado. Es el llamado período de contracción isovolumétrica o isométrica, lo que quiere decir que se produce aumento de la tensión en el músculo cardíaco, pero con un acortamiento escaso o nulo de las fibras musculares. Período de eyección Cuando la presión ventricular izquierda aumenta ligeramente por encima de 80 mmHg (y la presión ventricular derecha ligeramente por encima de 8 mmHg), las presiones ventriculares abren las válvulas semilunares. Inmediatamente comienza a salir la sangre de los ventrículos. Aproximadamente el 60% de la sangre del ventrículo al final de la diástole es expulsada durante la sístole; en torno al 70% de esta porción es expulsado durante el primer tercio del período de eyección y el 30% restante del vaciado se produce durante los dos tercios siguientes. Por tanto, el primer tercio se denomina período de eyección rápida y los dos tercios finales período de eyección lenta. Período de relajación isovolumétrica (isométrica) Al final de la sístole comienza súbitamente la relajación ventricular, lo que permite que las presiones intraventriculares derecha e izquierda disminuyan rápidamente. Las presiones elevadas de las grandes arterias distendidas que se acaban de llenar con la sangre que procede de los ventrículos que se han contraído empujan inmediatamente la sangre de nuevo hacia los ventrículos, lo que cierra súbitamente las válvulas aórtica y pulmonar. Durante otros 0,03 a 0,06 s el músculo cardíaco sigue relajándose, aun cuando no se modifica el volumen ventricular, dando lugar al período de relajación isovolumétrica o isométrica. Durante este período las presiones intraventriculares disminuyen rápidamente y regresan a sus bajos valores diastólicos. Después se abren las válvulas AV para comenzar un nuevo ciclo de bombeo ventricular.
Volumen telediastólico, volumen telesistólico y volumen sistólico Durante la diástole, el llenado normal de los ventrículos aumenta el volumen de cada uno de los ventrículos hasta aproximadamente 110 a 120 ml. Este volumen se denomina volumen telediastólico. Después, a medida que los ventrículos se vacían durante la sístole, el volumen disminuye aproximadamente 70 ml, lo que se denomina volumen sistólico. El volumen restante que queda en cada uno de los ventrículos, aproximadamente 40 a 50 ml, se denomina volumen telesistólico. La fracción del volumen telediastólico que es propulsada se denomina fracción de eyección, que habitualmente es igual a 0,6 (o el 60%) aproximadamente. Cuando el corazón se contrae con fuerza el volumen telesistólico puede disminuir hasta un valor tan bajo como 10 a 20 ml. Por el contrario, cuando fluyen grandes cantidades de sangre hacia los ventrículos durante la diástole, los volúmenes telediastólicos ventriculares pueden llegar a ser tan grandes como 150 a 180 ml en el corazón sano. Mediante el aumento del volumen telediastólico y la reducción del volumen telesistólico se puede aumentar el volumen sistólico hasta más del doble de lo normal. Las válvulas cardíacas evitan el flujo inverso de la sangre durante la sístole Válvulas auriculoventriculares Las válvulas AV (las válvulas tricúspide y mitral) impiden el flujo retrógrado de sangre desde los ventrículos hacia las aurículas durante la sístole, y las válvulas semilunares (es decir, las válvulas aórtica y de la arteria pulmonar) impiden el flujo retrógrado desde las arterias aorta y pulmonar hacia los ventrículos durante la diástole. Estas válvulas para el ventrículo izquierdo, se cierran y abren pasivamente. Es decir, se cierran cuando un gradiente de presión retrógrada empuja la sangre hacia atrás, y se abren cuando un gradiente de presión anterógrada fuerza la sangre en dirección anterógrada. Por motivos anatómicos, las válvulas AV, que están formadas por una película delgada, casi no precisan ningún flujo retrógrado para cerrarse, mientras que las válvulas semilunares, que son mucho más fuertes, precisan un flujo retrógrado bastante rápido durante algunos milisegundos.
Las válvulas cardíacas evitan el flujo inverso de la sangre durante la sístole «Válvulas auriculoventriculares« Las válvulas AV (las válvulas tricúspide y mitral) impiden el flujo retrógrado de sangre desde los ventrículos hacia las aurículas durante la sístole, y las válvulas semilunares (es decir, las válvulas aórtica y de la arteria pulmonar) impiden el flujo retrógrado desde las arterias aorta y pulmonar hacia los ventrículos durante la diástole.
Función de los músculos papilares los músculos papilares que se unen a los velos de las válvulas AV mediante las cuerdas tendinosas. Los músculos papilares se contraen cuando se contraen las paredes ventriculares, pero, al contrario de lo que se podría esperar, no contribuyen al cierre de las válvulas. Por el contrario, tiran de los velos de las válvulas hacia dentro, hacia los ventrículos, para impedir que protruyan demasiado hacia las aurículas durante la contracción ventricular. Si se produce la rotura de una cuerda tendinosa o si se produce parálisis de uno de los músculos papilares, la válvula protruye mucho hacia las aurículas durante la contracción ventricular, a veces tanto que se produce una fuga grave y da lugar a una insuficiencia cardíaca grave o incluso mortal. Válvulas aórtica y de la arteria pulmonar Las válvulas semilunares aórtica y pulmonar funcionan de una manera bastante distinta de las válvulas AV. Primero, las elevadas presiones de las arterias al final de la sístole hacen que las válvulas semilunares se cierren súbitamente, a diferencia del cierre mucho más suave de las válvulas AV. Segundo, debido a sus orificios más pequeños, la velocidad de la eyección de la sangre a través de las válvulas aórtica y pulmonar es mucho mayor que a través de las válvulas AV, que son mucho mayores. Además, debido al cierre rápido y a la eyección rápida, los bordes de las válvulas aórtica y pulmonar están sometidos a una abrasión mecánica mucho mayor que las válvulas AV. Las válvulas AV tienen el soporte de las cuerdas tendinosas, lo que no ocurre en el caso de las válvulas semilunares. A partir de la anatomía de las válvulas aórtica y pulmonar es evidente que deben estar situadas sobre una base de un tejido fibroso especialmente fuerte, pero muy flexible para soportar las tensiones físicas adicionales. Curva de presión aórtica Cuando el ventrículo izquierdo se contrae, la presión ventricular aumenta rápidamente hasta que se abre la válvula aórtica. Posteriormente, después de que se haya abierto la válvula, la presión del ventrículo aumenta mucho menos rápidamente, porque la sangre sale inmediatamente del ventrículo hacia la aorta y después hacia las arterias de distribución sistémica. La entrada de sangre en las arterias durante la sístole hace que sus paredes se distiendan y que la presión aumente hasta aproximadamente 120 mmHg. Al final de la sístole, después de que el ventrículo izquierdo haya dejado de impulsar sangre y se haya cerrado la válvula aórtica, las paredes elásticas de las arterias mantienen una presión elevada en las arterias, incluso durante la diástole. Se produce una incisura en la curva de presión aórtica cuando se cierra la válvula aórtica; está producida por un corto período de flujo retrógrado de sangre inmediatamente antes del cierre de la válvula, seguido por la interrupción súbita del flujo retrógrado. Después de que se haya cerrado la válvula aórtica, la presión en el interior de la aorta disminuye lentamente durante toda la sístole porque la sangre que está almacenada en las arterias elásticas distendidas fluye continuamente a través de los vasos periféricos de nuevo hacia las venas. Antes de que se contraiga de nuevo el ventrículo, la presión aórtica habitualmente ha disminuido hasta aproximadamente 80 mmHg (presión diastólica), que es dos tercios de la presión máxima de 120 mmHg (presión sistólica) que se produce en la aorta durante la contracción ventricular. Las curvas de presión del ventrículo derecho y de la arteria pulmonar son similares a las de la aorta, excepto que las presiones tienen una magnitud de solo aproximadamente 1/6.
Relación de los tonos cardíacos con el bombeo cardíaco Cuando se ausculta el corazón con un estetoscopio no se oye la apertura de las válvulas porque este es un proceso relativamente lento que no suele hacer ruido. Sin embargo, cuando las válvulas se cierran, los velos de las válvulas y los líquidos circundantes vibran bajo la influencia de los cambios súbitos de presión, generando un sonido que viaja en todas las direcciones a través del tórax. Cuando se contraen los ventrículos primero se oye un ruido que está producido por el cierre de las válvulas AV. El tono de la vibración es bajo y relativamente prolongado, y se conoce como el primer tono cardíaco. Cuando se cierran las válvulas aórtica y pulmonar al final de la sístole se oye un golpe seco y rápido porque estas válvulas se cierran rápidamente, y los líquidos circundantes vibran durante un período corto. Este sonido se denomina segundo tono cardíaco. Las causas precisas de los tonos cardíacos. Generación de trabajo del corazón El trabajo sistólico del corazón es la cantidad de energía que el corazón convierte en trabajo durante cada latido cardíaco mientras bombea sangre hacia las arterias. El trabajo minuto es la cantidad total de energía que se convierte en trabajo en 1 min; este parámetro es igual al trabajo sistólico multiplicado por la frecuencia cardíaca por minuto. El trabajo del corazón se utiliza de dos maneras. Primero, la mayor proporción se utiliza, con mucho, para mover la sangre desde las venas de baja presión hacia las arterias de alta presión. Esto se denomina trabajo volumen-presión o trabajo externo. Segundo, una pequeña proporción de la energía se utiliza para acelerar la sangre hasta su velocidad de eyección a través de las válvulas aórtica y pulmonar, que es el componente de energía cinética del flujo sanguíneo del trabajo cardíaco. El trabajo externo del ventrículo derecho es normalmente de alrededor de la sexta parte del trabajo del ventrículo izquierdo debido a la diferencia de seis veces de las presiones sistólicas que bombean los dos ventrículos. El trabajo adicional de cada uno de los ventrículos necesario para generar la energía cinética del flujo sanguíneo es proporcional a la masa de sangre que se expulsa multiplicada por el cuadrado de la velocidad de eyección. Habitualmente el trabajo del ventrículo izquierdo necesario para crear la energía cinética del flujo sanguíneo es de solo un 1% del trabajo total del ventrículo y, por tanto, se ignora en el cálculo del trabajo sistólico total. Sin embargo, en ciertas situaciones anormales, como en la estenosis aórtica, en la que la sangre fluye con una gran velocidad a través de la válvula estenosada, puede ser necesario más del 50% del trabajo total para generar la energía cinética del flujo sanguíneo.
La curva de presión diastólica se determina llenando el corazón con volúmenes de sangre progresivamente mayores y midiendo la presión diastólica inmediatamente antes de que se produzca la contracción ventricular, que es la presión telediastólica del ventrículo. La curva de presión sistólica se determina registrando la presión sistólica que se alcanza durante la contracción ventricular a cada volumen de llenado. Hasta que el volumen del ventrículo que no se está contrayendo no aumenta por encima de aproximadamente 150 ml, la presión «diastólica» no aumenta mucho. Por tanto, hasta este volumen la sangre puede fluir con facilidad hacia el ventrículo desde la aurícula. Por encima de 150 ml la presión diastólica ventricular aumenta rápidamente, en parte porque el tejido fibroso del corazón ya no se puede distender más y en parte porque el pericardio que rodea el corazón se ha llenado casi hasta su límite. Durante la contracción ventricular, la presión «sistólica» aumenta incluso a volúmenes ventriculares bajos y alcanza un máximo a un volumen ventricular de 150 a 170 ml. Después, a medida que sigue aumentando el volumen, la presión sistólica llega a disminuir en algunas situaciones, como se muestra por la disminución de la curva de presión sistólica porque a estos volúmenes elevados los filamentos de actina y de miosina de las fibras musculares cardíacas están tan separados que la fuerza de la contracción de cada una de las fibras cardíacas se hace menos óptima. Obsérvese especialmente en la figura que la presión sistólica máxima del ventrículo izquierdo normal está entre 250 y 300 mmHg, aunque esto varía mucho con la fuerza del corazón de cada persona y con el grado de la estimulación del corazón por los nervios cardíacos. Para el ventrículo derecho normal la presión sistólica máxima está entre 60 y 80 mmHg.
Conceptos de precarga y poscarga Cuando se evalúan las propiedades contráctiles del músculo es importante especificar el grado de tensión del músculo cuando comienza a contraerse, que se denomina precarga, y especificar la carga contra la que el músculo ejerce su fuerza contráctil, que se denomina poscarga. Para la contracción cardíaca habitualmente se considera que la precarga es la presión telediastólica cuando el ventrículo ya se ha llenado. La poscarga del ventrículo es la presión de la aorta que sale del ventrículo.
Regulación del bombeo cardíaco Cuando una persona está en reposo el corazón solo bombea de 4 a 6 l de sangre cada minuto. Durante el ejercicio intenso puede ser necesario que el corazón bombee de cuatro a siete veces esta cantidad. Los mecanismos básicos mediante los que se regula el volumen que bombea el corazón son: 1) regulación cardíaca intrínseca del bombeo en respuesta a los cambios del volumen de la sangre que fluye hacia el corazón 2) control de la frecuencia cardíaca y del bombeo cardíaco por el sistema nervioso autónomo.
Curvas de función ventricular Una de las mejores formas de expresar la capacidad funcional de los ventrículos de bombear sangre es mediante la curvas de función ventricular. Así, las curvas de función ventricular son otra forma de expresar el mecanismo de Frank-Starling del corazón. Es decir, a medida que los ventrículos se llenan en respuesta a unas presiones auriculares más altas, se produce aumento del volumen de los dos ventrículos y de la fuerza de la contracción del músculo cardíaco, lo que hace que el corazón bombee mayores cantidades de sangre hacia las arterias
Control del corazón por los nervios simpáticos y parasimpáticos La eficacia de la función de bomba del corazón también está controlada por los nervios simpáticos y parasimpáticos (vagos), que inervan de forma abundante el corazón. Para niveles dados de presión auricular de entrada, la cantidad de sangre que se bombea cada minuto (gasto cardíaco) con frecuencia se puede aumentar más de un 100% por la estimulación simpática. Por el contrario, el gasto se puede disminuir hasta un valor tan bajo como cero o casi cero por la estimulación vagal (parasimpática).
Mecanismos de excitación del corazón por los nervios simpáticos La estimulación simpática intensa puede aumentar la frecuencia cardíaca en seres humanos adultos jóvenes desde la frecuencia normal de 70 latidos/min hasta 180 a 200 y, raras veces, incluso 250 latidos/min. Además, la estimulación simpática aumenta la fuerza de la contracción cardíaca hasta el doble de lo normal, aumentando de esta manera el volumen de sangre que se bombea y aumentando la presión de eyección. Así, con frecuencia la estimulación simpática puede aumentar el gasto cardíaco máximo hasta dos o tres veces, además del aumento del gasto que produce el mecanismo de Frank-Starling que ya se ha comentado. Por el contrario, la inhibición de los nervios simpáticos del corazón puede disminuir la función de bomba del corazón en un grado moderado: en condiciones normales, las fibras nerviosas simpáticas que llegan al corazón descargan continuamente a una frecuencia baja que mantiene el bombeo aproximadamente un 30% por encima del que habría sin estimulación simpática. Por tanto, cuando la actividad del sistema nervioso simpático disminuye por debajo de lo normal, tanto de la frecuencia cardíaca como la fuerza de la contracción del músculo ventricular se reducen, con lo que disminuye el nivel de bombeo cardíaco hasta un 30% por debajo de lo normal. La estimulación parasimpática (vagal) reduce la frecuencia cardíaca y la fuerza de la contracción La estimulación intensa de las fibras nerviosas parasimpáticas de los nervios vagos que llegan al corazón puede interrumpir el latido cardíaco durante algunos segundos, pero después el corazón habitualmente «escapa» y late a una frecuencia de 20 a 40 latidos/min mientras continúe la estimulación parasimpática. Además, la estimulación vagal intensa puede reducir la fuerza de la contracción del músculo cardíaco en un 20-30%. Las fibras vagales se distribuyen principalmente por las aurículas y no mucho en los ventrículos, en los que se produce la contracción de potencia del corazón. Esta distribución explica por qué el efecto de la estimulación vagal tiene lugar principalmente sobre la reducción de la frecuencia cardíaca, en lugar de reducir mucho la fuerza de la contracción del corazón. Sin embargo, la gran disminución de la frecuencia cardíaca, combinada con una ligera reducción de la fuerza de la contracción cardíaca, puede reducir el bombeo ventricular en un 50% o más.
Efecto de los iones potasio y calcio sobre la función cardíaca Los iones potasio tienen un efecto marcado sobre los potenciales de membrana, los iones calcio desempeñan una función especialmente importante en la activación del proceso contráctil del músculo. Por tanto, cabe esperar que la concentración de cada uno de estos dos iones en los líquidos extracelulares también tenga efectos destacados sobre la función de bomba del corazón.
Efecto de los iones potasio El exceso de potasio hace que el corazón esté dilatado y flácido, y también reduce la frecuencia cardíaca. Grandes cantidades de potasio también pueden bloquear la conducción del impulso cardíaco desde las aurículas hacia los ventrículos a través del haz AV. La elevación de la concentración de potasio hasta solo 8 a 12 mEq/l (dos a tres veces el valor normal) puede producir una profunda debilidad del corazón, una alteración del ritmo e incluso la muerte. Estos efectos se deben parcialmente al hecho de que una concentración elevada de potasio en los líquidos extracelulares reduce el potencial de membrana en reposo de las fibras del músculo cardíaco. Es decir, la alta concentración de potasio en el líquido extracelular despolariza parcialmente la membrana celular, lo que provoca que el potencial de membrana sea menos negativo. Cuando disminuye el potencial de membrana también lo hace la intensidad del potencial de acción, lo que hace que la contracción del corazón sea progresivamente más débil. Efecto de los iones calcio Un exceso de iones calcio produce efectos casi exactamente contrarios a los de los iones potasio, haciendo que el corazón progrese hacia una contracción espástica. Este efecto está producido por el efecto directo de los iones calcio en el inicio del proceso contráctil cardíaco, como se explicó antes en este mismo capítulo. Por el contrario, el déficit de iones calcio produce debilidad cardíaca, similar al efecto de la elevación de la concentración de potasio. Afortunadamente las concentraciones de iones calcio en la sangre normalmente están reguladas en un intervalo muy estrecho. Por tanto, los efectos cardíacos de las concentraciones anormales de calcio raras veces tienen significado clínico. Efecto de la temperatura sobre la función cardíaca El aumento de la temperatura corporal, como ocurre durante la fiebre, produce un gran aumento de la frecuencia cardíaca, a veces hasta del doble del valor normal. El descenso de la temperatura produce una gran disminución de la frecuencia cardíaca, que puede disminuir hasta solo algunos latidos por minuto cuando una persona está cerca de la muerte por hipotermia en el intervalo de temperatura corporal de 16 °C a 21 °C. Estos efectos probablemente se deben al hecho de que el calor aumenta la permeabilidad de la membrana del músculo cardíaco a los iones que controlan la frecuencia cardíaca, acelerando el proceso de autoexcitación. La fuerza contráctil del corazón con frecuencia se incrementa transitoriamente cuando hay un aumento moderado de la temperatura, como ocurre durante el ejercicio corporal, aunque una elevación prolongada de la temperatura agota los sistemas metabólicos del corazón y finalmente produce debilidad. Por tanto, la función óptima del corazón depende mucho del control adecuado de la temperatura corporal mediante los mecanismos. El incremento de la carga de presión arterial (hasta un límite) no disminuye el gasto cardíaco , el aumento de la presión arterial en la aorta no reduce el gasto cardíaco hasta que la presión arterial media aumenta por encima de aproximadamente 160 mmHg. En otras palabras, durante la función normal del corazón a presiones arteriales sistólicas normales (80 a 140 mmHg) el gasto cardíaco está determinado casi totalmente por la facilidad del flujo sanguíneo a través de los tejidos corporales, que a su vez controla el retorno venoso de la sangre hacia el corazón.
Los sentidos del gusto y el olfato nos permiten distinguir los alimentos indeseables o incluso mortales de aquellos otros que resultan agradables de comer y nutritivos. Además, desencadenan respuestas fisiológicas que intervienen en la digestión y en la utilización de los alimentos. El sentido del olfato también permite que los animales reconozcan la proximidad de otros animales o hasta de cada individuo entre sus congéneres. Por último, ambos sentidos se encuentran íntimamente ligados a funciones emocionales y conductuales primitivas de nuestro sistema nervioso. En este capítulo, hablaremos de cómo se detectan los estímulos del gusto y el olfato y del modo en que se codifican en señales nerviosas transmitidas al encéfalo.
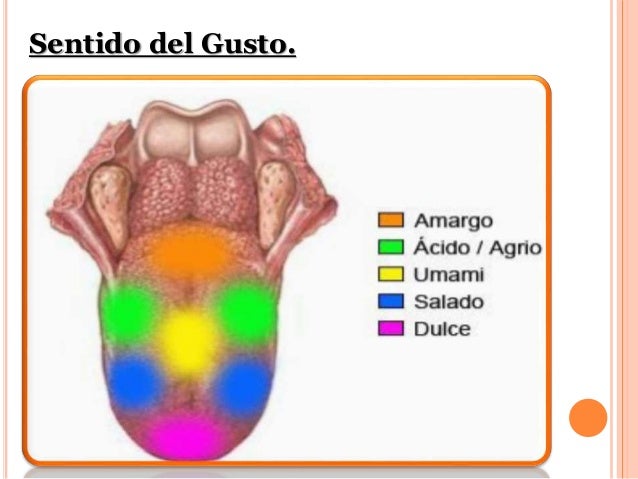
Sentido del gusto El gusto constituye sobre todo una función de las yemas gustativas de la boca, pero es una experiencia frecuente que el sentido del olfato también contribuya poderosamente a su percepción. Además, la textura de los alimentos, detectada por la sensibilidad táctil de la boca, y la presencia de sustancias que estimulen las terminaciones para el dolor, como la pimienta, modifica enormemente la experiencia gustativa. La importancia del gusto radica en el hecho de que permite a una persona escoger la comida en función de sus deseos y a menudo según las necesidades metabólicas de los tejidos corporales para cada sustancia específica. Sensaciones gustativas primarias No se conoce la identidad de todas las sustancias químicas específicas que excitan los diversos receptores gustativos. Los estudios psicofisiológicos y neurofisiológicos han identificado un mínimo de 13 receptores químicos probables en las células gustativas, de los siguientes tipos: 2 receptores para el sodio, 2 para el potasio, 1 para el cloruro, 1 para la adenosina, 1 para la inosina, 2 para el sabor dulce, 2 para el sabor amargo, 1 para el glutamato y 1 para el ion hidrógeno. Con el fin de realizar un análisis práctico del gusto, las capacidades señaladas de los receptores también se han reunido en cinco categorías generales llamadas sensaciones gustativas primarias. Estas son agrio, salado, dulce, amargo y «umami».
Sabor agrio El sabor agrio está causado por los ácidos, es decir, por la concentración del ion hidrógeno, y la intensidad de esta sensación gustativa es aproximadamente proporcional al logaritmo de esta concentración del ion hidrógeno (es decir, cuanto más ácido sea un alimento, más potente se vuelve dicha sensación). Sabor salado El sabor salado se despierta por las sales ionizadas, especialmente por la concentración del ion sodio. La cualidad de este rasgo varía de una sal a otra, porque algunas de ellas suscitan otras sensaciones gustativas además del sabor salado. Los cationes de las sales, sobre todo los cationes sodio, son los principales responsables del gusto salado, pero los aniones también contribuyen en menor medida. Sabor dulce El sabor dulce no está ocasionado por una sola clase de sustancias químicas. Entre los tipos de productos que lo originan figuran los azúcares, glicoles, alcoholes, aldehídos, cuerpos cetónicos, amidas, ésteres, ciertos aminoácidos, algunas proteínas pequeñas, los ácidos sulfónicos, los ácidos halogenados y las sales inorgánicas de plomo y berilio. Obsérvese en concreto que la mayoría de las sustancias que generan el sabor dulce son compuestos orgánicos. Resulta especialmente interesante que unas ligeras modificaciones en la estructura química, como la incorporación de un simple radical, muchas veces pueden cambiar el producto de dulce a amargo. Sabor amargo El sabor amargo, igual que el sabor dulce, no está originado por un único tipo de agente químico. En este caso, una vez más las sustancias que lo suministran son casi todas orgánicas. Dos clases particulares tienen una especial probabilidad de causar sensaciones de sabor amargo: 1) las sustancias orgánicas de cadena larga que contienen nitrógeno, y 2) los alcaloides. Estos últimos comprenden muchos de los fármacos empleados en medicamentos como la quinina, la cafeína, la estricnina y la nicotina. Algunas sustancias que al principio saben saladas dejan un regusto amargo. Esta característica sucede con la sacarina, lo que le otorga un carácter desagradable para algunas personas. El sabor amargo, cuando se da con una gran intensidad, suele hacer que la persona o el animal rechace la comida. Esta reacción es una función indudablemente importante de dicha sensación gustativa, pues muchas toxinas mortales presentes en las plantas venenosas son alcaloides, y prácticamente todos estos alcaloides suscitan un sabor amargo intenso, normalmente seguido por el rechazo del alimento. Sabor umami Umami, una palabra japonesa que significa «delicioso», designa una sensación gustativa agradable que resulta diferente desde el punto de vista cualitativo de los sabores agrio, salado, dulce o amargo.Umami es el sabor dominante de los alimentos que contienen l-glutamato, como los extractos cárnicos y el queso curado, y algunos fisiólogos lo consideran una quinta categoría independiente de estímulos gustativos primarios. Un receptor gustativo para el l-glutamato puede estar relacionado con uno de los receptores glutamatérgicos expresado también en las sinapsis neuronales del cerebro. Sin embargo, aún no están claros los mecanismos moleculares exactos responsables del sabor umami.
Umbral gustativo El umbral de estimulación para el sabor agrio debido al ácido clorhídrico oscila alrededor de 0,0009 M; en el caso del sabor salado por el cloruro sódico es de 0,01 M; para el sabor dulce por la sacarosa es de 0,01 M, y para el sabor amargo por la quinina, de 0,000008 M. Obsérvese sobre todo la mayor sensibilidad para las sensaciones gustativas amargas que para todas las demás, lo que ya resultaba previsible, pues esta sensación cumple una función protectora importante contra muchas toxinas peligrosas de los alimentos. Ceguera gustativa Algunas personas están ciegas para el gusto de ciertas sustancias, sobre todo los diversos tipos de compuestos de la tiourea. Un producto empleado a menudo por parte de los psicólogos para poner de manifiesto la ceguera gustativa es la feniltiocarbamida, para la que de un 15 a un 30% de las personas muestran una ceguera gustativa; el porcentaje exacto depende del método de exploración y de la concentración de la sustancia. Yemas gustativas y su función Entretejida alrededor de los cuerpos de las células gustativas hay toda una red terminal ramificada de fibras nerviosas gustativas que reciben el estímulo de las células receptoras del gusto. Algunas se invaginan en pliegues de la membrana de la célula gustativa. Debajo de la membrana celular se forman muchas vesículas cerca de las fibras. Se cree que estas vesículas contienen una sustancia neurotransmisora que se libera a través de la membrana celular para excitar las terminaciones de las fibras nerviosas como respuesta a la estimulación gustativa.
Las yemas gustativas se encuentran en los tres tipos siguientes de papilas linguales: 1) una gran cantidad está en las paredes de las depresiones que rodean a las papilas caliciformes, que forman una línea en «V» sobre la superficie de la parte posterior de la lengua; 2) un número moderado queda sobre las papilas fungiformes en la cara anterior plana de la lengua, y 3) una proporción también moderada se encuentra sobre las papilas foliáceas situadas en los pliegues a lo largo de las superficies laterales de la lengua. Existen otras yemas gustativas más en el paladar, y unas pocas en los pilares amigdalinos, en la epiglotis e incluso en la parte proximal del esófago. Los adultos poseen de 3.000 a 10.000 yemas gustativas y los niños tienen unas pocas más. Pasados los 45 años, muchas yemas degeneran, lo que deriva en que la sensibilidad del gusto disminuya en el anciano La membrana de la célula gustativa, igual que la mayoría de las demás células receptoras sensitivas, tiene una carga negativa en su interior con respecto al exterior. La aplicación de una sustancia con sabor sobre los cilios gustativos provoca una pérdida parcial de este potencial negativo, es decir, la célula gustativa se despolariza. En la mayoría de los casos, el descenso del potencial, dentro de un rango amplio, es aproximadamente proporcional al logaritmo de la concentración de la sustancia estimulante. Este cambio del potencial eléctrico en la célula gustativa se llama potencial de receptor para el gusto. Tras la primera aplicación del estímulo gustativo, la frecuencia de descarga de las fibras nerviosas procedentes de las yemas gustativas asciende hasta un máximo en una pequeña fracción de segundo, pero a continuación se adapta durante los segundos siguientes hasta regresar a un nivel estable más bajo mientras permanezca presente el estímulo gustativo. Por tanto, el nervio gustativo transmite una señal potente inmediata, y una señal continua más débil todo el tiempo que la yema gustativa siga expuesta al estímulo correspondiente.
Transmisión de las señales gustativas en el sistema nervioso central. Los impulsos gustativos procedentes de los dos tercios anteriores de la lengua se dirigen primero hacia el nervio lingual, a continuación van por la cuerda del tímpano hacia el nervio facial, y finalmente llegan al tracto solitario en el tronco del encéfalo. Las sensaciones gustativas de las papilas caliciformes situadas en el dorso de la lengua y en otras regiones posteriores de la boca y de la garganta se transmiten a través del nervio glosofaríngeo también hacia el tracto solitario, pero a un nivel un poco más inferior. Finalmente, unas cuantas señales gustativas se conducen hacia el tracto solitario desde la base de la lengua y otras porciones de la región faríngea por medio del nervio vago.
Integración de los reflejos gustativos en el tronco del encéfalo Desde el tracto solitario, muchas señales gustativas se transmiten directamente por el propio tronco del encéfalo hacia los núcleos salivales superior e inferior, y estas zonas envían señales hacia las glándulas submandibular, sublingual y parótida que sirven para controlar la secreción de saliva durante la ingestión y la digestión de la comida. Rápida adaptación del gusto Todo el mundo está acostumbrado al hecho de que las sensaciones gustativas se adaptan con rapidez; muchas veces lo hacen prácticamente por completo en un plazo de 1 min más o menos tras su estimulación continua. Con todo, según los estudios electrofisiológicos realizados con las fibras nerviosas gustativas, está claro que la adaptación de las propias yemas gustativas normalmente no explica más que la mitad de esta rápida adaptación del gusto. Por tanto, el grado final de adaptación tan extremo que sucede en el sentido del gusto ocurre casi con seguridad en el sistema nervioso central, aunque no se conozcan cuáles son sus mecanismos. En cualquier caso, se trata de un fenómeno diferente del que se da en la mayoría de los demás sistemas sensitivos, cuya adaptación se produce principalmente a nivel de los receptores. Preferencias gustativas y control del régimen alimentario Las preferencias gustativas no significan nada más que un animal elegirá ciertos tipos de comida por encima de otros, y que recurre automáticamente a este mecanismo como medio para controlar el tipo de alimentación que consume. Además, sus preferencias gustativas cambian a menudo en función de las necesidades corporales de ciertas sustancias específicas. Los siguientes experimentos ponen de manifiesto esta capacidad de los animales para escoger la comida según las necesidades de sus organismos. En primer lugar, después de una suprarrenalectomía los animales hiponatrémicos se decantan automáticamente por beber agua con una concentración elevada de cloruro sódico por encima del agua pura, y muchas veces la cantidad de cloruro de sodio en el agua basta para cubrir las necesidades corporales y evitar la muerte por pérdida de sodio. En segundo lugar, un animal que reciba inyecciones con una cantidad excesiva de insulina sufre una pérdida de azúcar en la sangre y selecciona mecánicamente la más dulce de las comidas entre muchas opciones. En tercer lugar, los animales paratiroidectomizados con pérdida de calcio se inclinan instintivamente por beber agua con una concentración elevada de cloruro cálcico. Estos mismos fenómenos también se observan en la vida cotidiana.
SENTIDO DEL OLFATO. El olfato es el menos conocido de nuestros sentidos, debido en parte al hecho de que constituye un fenómeno subjetivo que no puede estudiarse con facilidad en los animales inferiores. Otro problema que complica la situación es que el sentido del olfato está poco desarrollado en los seres humanos en comparación con lo que sucede en muchos animales inferiores. Membrana olfatoria La membrana olfatoria, En sentido medial, se dobla hacia abajo a lo largo de la superficie del tabique en su parte superior; en sentido lateral se pliega sobre el cornete superior e incluso sobre una pequeña porción de la cara superior del cornete medio. El compuesto oloroso se une a la porción de la proteína receptora que se vuelve hacia el exterior. Sin embargo, la parte interna de la proteína plegada está acoplada a la proteína G, que es en sí una combinación de tres subunidades. Al excitarse la proteína receptora se desprende una subunidad α de la proteína G y activa la adenilato ciclasa, que está fija al interior de la membrana ciliar cerca del cuerpo de la célula receptora. A su vez, la ciclasa activada convierte muchas moléculas de trifosfato de adenosina intracelular en monofosfato de adenosina cíclico (AMPc). Finalmente, este AMPc activa otra proteína cercana de la membrana, un canal activado para el ion sodio, que abre su «compuerta» y permite el vertido de una gran cantidad de iones sodio a través de la membrana hacia el citoplasma de la célula receptora. Los iones sodio elevan el potencial eléctrico dentro de la membrana celular en sentido positivo, lo que excita a la neurona olfatoria y transmite potenciales de acción hacia el sistema nervioso central por medio del nervio olfatorio.
ormemente el efecto excitador hasta del más débil de los compuestos olorosos. En resumen: 1) la activación de la proteína receptora por la sustancia olorosa estimula el complejo de la proteína G; 2) esto a su vez activa múltiples moléculas de adenilato ciclasa por dentro de la membrana de la célula olfatoria; 3) esto provoca la formación de un número muchas veces mayor de moléculas de AMPc, y, finalmente, 4) el AMPc abre una cantidad todavía muy superior de canales iónicos de sodio. Por tanto, incluso la concentración más minúscula de un producto oloroso específico pone en marcha un efecto en cascada que abre una proporción elevadísima de canales de sodio. Este proceso explica la exquisita sensibilidad de las neuronas olfatorias incluso frente a la cantidad más leve de sustancia olorosa. Además del mecanismo químico básico por el que se activan las células olfatorias, diversos factores físicos influyen sobre su grado de estimulación. En primer lugar, solo es posible oler las sustancias volátiles que pueden inhalarse por las narinas. En segundo lugar, la sustancia estimulante ha de tener al menos un carácter un poco hidrosoluble para que sea capaz de atravesar el moco y llegar a los cilios olfatorios. En tercer lugar, es útil que además sea como mínimo un tanto liposoluble, se supone que debido a que los componentes lipídicos del cilio constituyen una débil barrera para los productos que no sean liposolubles.
Potenciales de membrana y potenciales de acción en las células olfatorias El potencial de membrana en el interior de las células olfatorias sin estimular, según se recoge mediante microelectrodos, oscila alrededor de –55 mV. A este nivel, la mayor parte de las células generan potenciales de acción continuos a una frecuencia muy baja, que varía desde una vez cada 20s hasta dos a tres por segundo. La mayoría de las sustancias olorosas producen una despolarización de la membrana en la célula olfatoria, lo que disminuye el potencial negativo de la célula desde su valor normal de –55 mV hasta – 30 o menos aún: es decir, cambia el voltaje en un sentido positivo. Junto a esto, el número de potenciales de acción crece de 20 a 30 por segundo, lo que representa una frecuencia alta para las diminutas fibras nerviosas olfatorias. Dentro de un amplio intervalo, la frecuencia de impulsos del nervio olfatorio cambia aproximadamente en proporción al logaritmo de la intensidad del estímulo, lo que manifiesta que los receptores olfatorios obedecen a unos principios de transducción similares a los que siguen otros receptores sensitivos. Rápida adaptación de las sensaciones olfatorias. Los receptores olfatorios se adaptan alrededor del 50% más o menos durante el primer segundo después de su estimulación. A partir de ahí, el proceso ya sigue muy poco más y con una gran lentitud. En cambio, todos sabemos por nuestra propia experiencia que las sensaciones olfatorias se adaptan casi hasta su extinción en un plazo en torno a 1 min después de entrar en una atmósfera cargada con un olor muy penetrante. Como esta adaptación psicológica resulta mucho mayor que el grado de adaptación de los receptores, es casi seguro que la mayor parte del proceso suplementario sucede dentro del sistema nervioso central, lo que también parece ser así en el caso de la adaptación a las sensaciones gustativas. Indagación de las sensaciones olfatorias primarias En el pasado, la mayoría de los fisiólogos estaban convencidos de que muchas de las sensaciones olfatorias se encuentran a cargo de unas cuantas sensaciones primarias bastante independientes, de forma parecida a lo que sucede con la visión y el gusto, que derivan de unas pocas sensaciones primarias determinadas. A raíz de los estudios psicológicos, un intento de clasificar estas sensaciones es el siguiente: 1. Alcanforado. 2. Almizcleño. 3. Floral. 4. Mentolado. 5. Etéreo. 6. Acre. 7. Pútrido.
Naturaleza afectiva del olfato El olfato, aún más que el gusto, posee una cualidad afectiva agradable o desagradable, y por ello probablemente aún es más importante que este sentido en la selección de los alimentos. En efecto, una persona que haya consumido con anterioridad una comida que le sentara mal suele sentir náuseas ante su olor en una segunda ocasión. A la inversa, un perfume con las cualidades correctas puede ser un poderoso estimulante en las emociones humanas. Por ende, en algunos animales inferiores los olores cumplen la misión de excitantes primarios del impulso sexual. Umbral para el olfato Una de las principales características del olfato es la minúscula cantidad del agente estimulante presente en el aire que es capaz de suscitar una sensación olfatoria. Por ejemplo, la sustancia metilmercaptano puede olerse con la presencia solo de una 25 billonésima de gramo en cada mililitro de aire. Debido a este umbral tan bajo, dicha sustancia se mezcla con el gas natural para otorgarle un olor que pueda detectarse cuando se fugue una cantidad aún pequeña de una tubería. Gradaciones de las intensidades del olor Aunque las concentraciones umbrales de las sustancias que suscitan los olores son pequeñísimas, para muchos productos olorosos (si no para la mayoría), unos valores nada más que de 10 a 50 veces por encima del umbral provocan la máxima intensidad olfatoria. Este intervalo de discriminación de la intensidad choca con lo que sucede en la mayor parte de los demás sistemas sensitivos del cuerpo, cuyos límites entre los que se distinguen las intensidades son inmensos: por ejemplo, de 500.000 a 1 en el caso de los ojos y de 1 billón a 1 en el del oído.
Vías olfatorias primitivas y nuevas hacia el sistema nervioso central El tracto olfatorio penetra en el encéfalo a nivel de la unión anterior entre el mesencéfalo y el cerebro; allí, se divide en dos vías. El sistema olfatorio antiguo: el área olfatoria lateral El área olfatoria lateral está compuesta sobre todo por las cortezas prepiriforme y piriforme además de la porción cortical de los núcleos amigdalinos. Desde estas zonas, las vías activadoras se dirigen hacia casi todas las porciones del sistema límbico, en especial hacia las menos primitivas como el hipocampo, que parece más importante para aprender a disfrutar de ciertos alimentos o a aborrecerlos en función de las experiencias personales vividas con ellos. Por ejemplo, se cree que esta área olfatoria lateral y sus abundantes conexiones con el sistema límbico de orden conductual hacen que una persona desarrolle una absoluta aversión hacia las comidas que le hayan provocado náuseas y vómitos. La vía moderna Últimamente se ha descubierto una vía olfatoria más reciente que atraviesa el tálamo, pasando por su núcleo dorsomedial y llegando después al cuadrante lateroposterior de la corteza orbitofrontal. Según los estudios con monos, este sistema más moderno probablemente interviene en el análisis consciente de los olores. Control centrífugo de la actividad en el bulbo olfatorio por parte del sistema nervioso central Muchas fibras nerviosas que nacen en las porciones olfatorias del cerebro siguen un trayecto hacia el exterior por el tracto olfatorio hasta el bulbo olfatorio (es decir, «centrífugo» desde el cerebro hacia la periferia). Estas fibras nerviosas acaban en una gran cantidad de pequeños granos situados entre las células mitrales y en penacho en el bulbo olfatorio. Los granos envían señales inhibidoras hacia estos dos tipos de células. Se cree que esta retroalimentación negativa podría constituir un medio para acentuar la capacidad específica de distinguir un olor de otro.
La membrana timpánica y el sistema de huesecillos. Conducción del sonido desde la membrana timpánica hasta la cóclea . La membrana timpánica (llamada corrientemente tímpano) y los huesecillos, que conducen el sonido desde ella hasta la cóclea (el oído interno) a través del oído medio. En la membrana timpánica se fija el manubrio o mango del martillo. Este hueso está unido al yunque por unos ligamentos diminutos, por lo que cualquier movimiento del primero arrastra al segundo con él. El extremo opuesto del yunque se articula con la cabeza del estribo y la base de este último descansa sobre el laberinto membranoso de la cóclea en la abertura de la ventana oval.
El extremo final del manubrio del martillo se fija al centro de la membrana timpánica y sobre este punto de inserción tira constantemente el músculo tensor del tímpano, que mantiene tensa dicha estructura. Esta tensión permite que las vibraciones sonoras de cualquier porción de esta membrana se transmitan a los huesecillos, lo que no sucedería si se encontrara relajada. Los huesecillos del oído medio están suspendidos por ligamentos de un modo tal que el martillo y el yunque actúan en combinación como una sola palanca, cuyo fulcro queda aproximadamente en el margen de la membrana timpánica. La articulación del yunque con el estribo hace que este último: 1) empuje hacia delante la ventana oval y el líquido coclear que está presente al otro lado cada vez que la membrana timpánica se mueve hacia dentro, y 2) tire del líquido hacia atrás cada vez que el martillo se desplaza hacia fuera.
«Ajuste de impedancias» a cargo del sistema de huesecillos La amplitud de los movimientos de la base del estribo con cada vibración sonora no supone nada más que tres cuartas partes del recorrido que efectúa el manubrio del martillo. Por tanto, el sistema de palanca osicular no aumenta la distancia del desplazamiento del estribo, tal como se cree habitualmente. Por el contrario, lo que en realidad hace es reducirlo, pero incrementar la fuerza de empuje alrededor de 1,3 veces. Además, la superficie de la membrana timpánica mide un área de unos 55 mm2, mientras que la del estribo presenta una media de 3,2 mm2. Esta diferencia de 17 veces multiplicada por la proporción de 1,3 que corresponde al sistema de palanca hace que la fuerza total a la que está sometido el líquido coclear sea unas 22 veces mayor que la ejercida por las ondas sonoras sobre la membrana timpánica. Dado que el líquido posee una inercia mucho mayor que el aire, hace falta un grado superior de fuerza para ocasionar la vibración del primero. Así pues, la membrana timpánica y el sistema de huesecillos aportan un ajuste de impedancias entre las ondas sonoras del aire y las vibraciones sonoras en el líquido de la cóclea. En efecto, el ajuste de impedancias está alrededor del 50 al 75% de la situación ideal para las frecuencias sonoras entre 300 y 3.000 ciclos/s, lo que permite utilizar la mayor parte de la energía portada por las ondas sonoras entrantes. Si falta el sistema de huesecillos y la membrana timpánica, las ondas sonoras aún pueden viajar directamente a través del aire contenido en el oído medio y entrar en la cóclea por la ventana oval. Sin embargo, en estas circunstancias la sensibilidad auditiva es de 15 a 20 decibelios menor que para la transmisión osicular, lo que equivale a un descenso desde un nivel intermedio de voz hasta otro apenas perceptible. Atenuación del sonido mediante la contracción de los músculos estapedio y tensor del tímpano Cuando se transmiten sonidos fuertes a través del sistema de huesecillos y desde él al sistema nervioso central, se desencadena un reflejo pasado un período de latencia que solo dura de 40 a 80 ms y que provoca la contracción del músculo estapedio o del estribo y, en menor medida, del músculo tensor del tímpano. Este último tira del manubrio del martillo hacia dentro mientras que el primero tira del estribo hacia fuera. Ambas fuerzas se oponen entre sí y de ese modo hacen que el sistema de huesecillos adquiera en su conjunto una mayor rigidez, lo que disminuye mucho la conducción osicular de los sonidos de baja frecuencia, especialmente por debajo de 1.000 ciclos/s. Este reflejo de atenuación es capaz de reducir la intensidad de transmisión para los sonidos de baja frecuencia de 30 a 40 decibelios, que es más o menos la misma diferencia que existe entre una voz fuerte y un susurro. Se piensa que este mecanismo cumple una función doble: proteger la cóclea de las vibraciones lesivas ocasionadas por un sonido excesivamente fuerte y ocultar los sonidos de baja frecuencia en un ambiente ruidoso. La ocultación normalmente elimina un componente importante del ruido de fondo y permite que una persona se concentre en los sonidos por encima de 1.000 ciclos/s, que contienen la mayor parte de la información pertinente para la comunicación vocal. Otra función de los músculos estapedio y tensor del tímpano consiste en disminuir la sensibilidad auditiva de una persona hacia sus propias palabras. Este efecto es suscitado por unas señales nerviosas colaterales transmitidas hacia estos músculos al mismo tiempo que el cerebro activa el mecanismo de la voz. Transmisión del sonido a través del hueso Debido a que el oído interno, la cóclea o caracol, está enterrado en una cavidad ósea del hueso temporal, llamada laberinto óseo, las vibraciones sufridas por el cráneo en su conjunto pueden originar vibraciones en el líquido de la cóclea. Por tanto, en las condiciones adecuadas, un diapasón o un vibrador electrónico colocado sobre cualquier protuberancia ósea del cráneo, pero especialmente en la apófisis mastoides cercana al oído, hace que la persona escuche el sonido. Sin embargo, la energía que arrastra por el aire incluso un sonido fuerte no basta para causar la audición a través de la conducción ósea a no ser que se aplique un aparato electromecánico especial para la amplificación del sonido en el hueso.
Cóclea Anatomía funcional de la cóclea La cóclea es un sistema de tubos en espiral, consta de tres tubos enrollados uno junto a otro: 1) la rampa vestibular; 2) el conducto coclear o rampa media, y 3) la rampa timpánica. La rampa vestibular y el conducto coclear están separados por la membrana de Reissner (también llamada membrana vestibular). la rampa timpánica y el conducto coclear están divididos por la membrana o lámina basilar. Sobre su superficie se encuentra el órgano de Corti, que contiene una serie de células sensibles a estímulos electromecánicos, las células ciliadas. Se trata de los órganos receptores terminales que generan impulsos nerviosos como respuesta a las vibraciones sonoras.
La lámina basilar de la cóclea es una membrana fibrosa que separa el conducto coclear de la rampa timpánica. Contiene de 20.000 a 30.000 fibras basilares que se proyectan desde el centro óseo de la cóclea, el modíolo o columela, hacia su pared externa. Estas fibras son estructuras rígidas, elásticas, parecidas a lengüetas, que están fijas por su extremo basal al componente óseo central de la cóclea (el modíolo), pero esto no sucede en su extremo distal, donde solo se encuentran enterradas en la laxa estructura de la membrana. Dado que las fibras son rígidas y uno de sus extremos queda libre, pueden vibrar como las lengüetas de una armónica. La longitud de las fibras basilares aumenta progresivamente a partir de la ventana oval en sentido desde la base de la cóclea hacia su vértice o cúpula; las dimensiones pasan de unos 0,04 mm cerca de las ventanas oval y redonda hasta 0,5 mm en el extremo de la cóclea (el «helicotrema»), un cambio de 12 órdenes en su longitud. Sin embargo, el diámetro de las fibras disminuye desde la ventana oval hacia el helicotrema, por lo que su rigidez global desciende más de 100 veces. En consecuencia, las fibras cortas y rígidas cercanas a la ventana oval de la cóclea vibran mejor a una frecuencia muy alta, mientras que las fibras largas y flexibles próximas a su extremo final lo hacen mejor a una frecuencia baja. Así pues, la resonancia de las frecuencias altas en la lámina basilar se produce cerca de su base, zona por donde penetran las ondas sonoras en la cóclea a través de la ventana oval. Sin embargo, la resonancia de las frecuencias bajas sucede cerca del helicotrema, sobre todo debido a sus fibras menos rígidas, pero también por estar más «sobrecargadas» con un volumen de líquido extra que ha de vibrar a lo largo de los túbulos de la cóclea. El movimiento ascendente de la fibra basilar arrastra la membrana reticular hacia arriba y hacia dentro para acercarla al modíolo. A continuación, cuando la lámina basilar desciende, la membrana reticular se balancea hacia abajo y hacia fuera. El desplazamiento hacia dentro y hacia fuera hace que los cilios de las células ciliadas batan atrás y adelante contra la membrana tectoria. Así pues, las células ciliadas se excitan siempre que vibra la lámina basilar El órgano de corti es el órgano receptor que genera los impulsos nerviosos como respuesta a la vibración de la lámina basilar. Obsérvese su situación sobre la superficie de las fibras basilares y la lámina basilar. Los auténticos receptores sensitivos del órgano de Corti son dos tipos especializados de células nerviosas llamadas células ciliadas: una sola fila de células ciliadas internas, que suman unas 3.500 y poseen un diámetro de unos 12 μm, y tres o cuatro filas de células ciliadas externas, que totalizan alrededor de 12.000 y cuyo diámetro no mide nada más que alrededor de 8 micrómetros. La base y las caras laterales de las células ciliadas hacen sinapsis con una red de terminaciones nerviosas cocleares. Entre el 90 y el 95% de ellas acaban sobre las células ciliadas internas, lo que subraya su importancia especial para la detección del sonido.
Las fibras nerviosas estimuladas por las células ciliadas llegan al ganglio espiral de Corti, que está situado en el modíolo (el centro) de la cóclea.
Las señales auditivas se transmiten sobre todo por las células ciliadas internas Incluso aunque hay de tres a cuatro veces más células ciliadas externas que internas, aproximadamente el 90% de las fibras del nervio coclear son estimuladas por estas últimas en vez de por las primeras. No obstante, si se lesionan las células externas y las internas permanecen a pleno rendimiento, se produce una hipoacusia de grandes proporciones. Así las cosas, se ha propuesto que las células ciliadas externas controlan de algún modo la sensibilidad de las internas a los diferentes tonos de sonido, fenómeno denominado «ajuste» del sistema receptor. A favor de este concepto se postula el hecho de que es muy abundante el número de fibras nerviosas retrógradas que van desde el tronco del encéfalo hasta las inmediaciones de las células ciliadas externas. Su estimulación puede causar el acortamiento de las células ciliadas externas y tal vez modificar también su grado de rigidez. Estos efectos permiten pensar en un mecanismo nervioso retrógrado encargado de controlar la sensibilidad del oído a los diversos tonos sonoros, que esté activado por las células ciliadas externas. Potenciales de receptor de las células ciliadas y excitación de las fibras nerviosas auditivas Los estereocilios (es decir, los «cilios» que sobresalen desde los extremos de las células ciliadas) son estructuras duras debido a que poseen un armazón rígido de proteínas. Cada célula ciliada posee unos 100 estereocilios sobre su borde apical. Estos estereocilios van creciendo progresivamente hacia su lado más alejado del modíolo, y la parte superior de los estereocilios más cortos está sujeta por unos filamentos delgados a las porciones posteriores de los estereocilios vecinos más largos. Por tanto, cada vez que los cilios se inclinen en dirección hacia los más largos, tiran del extremo de los más pequeños hacia fuera desde la superficie de la célula ciliada. Esto provoca un fenómeno de transducción mecánica que abre de 200 a 300 canales de conducción catiónica, lo que permite el movimiento rápido de iones potasio con carga positiva desde el líquido del conducto coclear adyacente hacia los estereocilios, y esto suscita la despolarización de la membrana de la célula ciliada. Por tanto, cuando las fibras basilares se inclinan hacia la rampa vestibular, las células ciliadas se despolarizan, y cuando se mueven en el sentido opuesto se hiperpolarizan, lo que genera así un potencial de receptor alterno en su seno, lo que a su vez estimula las terminaciones del nervio coclear que hacen sinapsis en la base de las células ciliadas. Se cree que durante la despolarización las células ciliadas liberan un neurotransmisor de acción rápida en estas sinapsis. Es posible que la sustancia transmisora sea glutamato, pero no hay ninguna seguridad al respecto.
Potencial endococlear Para entender aún más a fondo los potenciales eléctricos generados por las células ciliadas, tenemos que explicar otro fenómeno eléctrico llamado potencial endococlear. El conducto coclear está ocupado por un líquido denominado endolinfa, a diferencia de la perilinfa presente en las rampas vestibular y timpánica. Estas dos últimas presentan una comunicación directa con el espacio subaracnoideo que rodea al encéfalo, de modo que la perilinfa es casi idéntica al líquido cefalorraquídeo. Por el contrario, la endolinfa que llena el conducto coclear es un líquido totalmente diferente de cuya secreción se encarga la estría vascular, una zona muy vascularizada situada en la pared externa de este conducto. La endolinfa contiene una concentración elevada de potasio y baja de sodio, situación que es exactamente la contraria a la composición de la perilinfa. Todo el tiempo existe un potencial eléctrico de unos +80 mV entre la endolinfa y la perilinfa, siendo positivo el interior del conducto coclear y negativo el exterior. Esto se llama potencial endococlear, y está generado por la secreción continua de iones potasio positivos hacia el conducto coclear por parte de la estría vascular.La importancia del potencial endococlear consiste en que la parte superior de las células ciliadas está proyectada hacia la membrana reticular y queda sumergida en la endolinfa del conducto coclear, mientras que la perilinfa baña su cuerpo situado en la parte inferior de la célula. Por ende, las células ciliadas poseen un potencial intracelular negativo de – 70 mV con respecto a la perilinfa, pero de –150 mV con respecto a la endolinfa en sus caras superiores, donde los cilios se proyectan a través de la membrana reticular hacia esta última. Se cree que dicho potencial eléctrico elevado en la punta de los estereocilios sensibiliza un grado más a la célula, lo que incrementa su capacidad para responder a los sonidos más tenues. Determinación de la frecuencia del sonido: el principio de la «posición» Si se parte de las explicaciones anteriores, resulta patente que los sonidos de baja frecuencia dan lugar a una activación máxima de la lámina basilar cerca de la cúpula de la cóclea, y los de alta frecuencia lo hacen cerca de su base. Los sonidos de una frecuencia intermedia activan la membrana a una distancia también intermedia entre ambos extremos. Por añadidura, las fibras nerviosas presentan una organización espacial dentro de la vía coclear, que se conserva durante todo el trayecto desde la cóclea hasta la corteza cerebral. El registro de señales en los fascículos auditivos del tronco del encéfalo y en los campos receptores auditivos de la corteza cerebral muestra que cada frecuencia sonora específica activa unas neuronas concretas del encéfalo. Por tanto, el método fundamental empleado por el sistema nervioso para detectar las diversas frecuencias sonoras consiste en determinar el punto más estimulado a lo largo de la lámina basilar, que se denomina principio de la posición para la determinación de la frecuencia sonora.
Determinación del volumen El sistema auditivo determina el volumen recurriendo a tres procedimientos como mínimo. En primer lugar, según sube el volumen sonoro, también aumenta la amplitud de la vibración en la lámina basilar y en las células ciliadas, por lo que estas últimas excitan las terminaciones nerviosas a una frecuencia más rápida. En segundo lugar, a medida que aumenta la amplitud de la vibración, hace que se estimule un número cada vez mayor de células ciliadas en la periferia de la porción resonante de la lámina basilar, lo que da lugar a una sumación espacial de los impulsos: es decir, la transmisión a través de muchas fibras nerviosas en vez de solo unas pocas. En tercer lugar, las células ciliadas externas no se estimulan apreciablemente hasta que la vibración de la lámina basilar alcanza una intensidad elevada y la activación de tales células probablemente comunica al sistema nervioso la información de que el sonido es fuerte. La unidad del decibelio Debido a los cambios extremos en las intensidades sonoras que el oído es capaz de detectar y distinguir, esta variable suele expresarse en forma del logaritmo de su intensidad real. Un aumento de 10 veces en la energía del sonido se denomina 1 belio, y 0,1 belios reciben el nombre de 1 decibelio. Un decibelio representa un incremento real de 1,26 veces en la energía sonora. Otra razón para emplear el sistema de decibelios en la expresión de las variaciones de volumen estriba en que, dentro del intervalo habitual de intensidades sonoras utilizado para la comunicación, el oído apenas es capaz de distinguir un cambio aproximado de 1 decibelio en esta variable. Mecanismos auditivos centrales Vías nerviosas auditivas Desde esta estructura, la vía auditiva asciende a través del lemnisco lateral. Parte de las fibras acaban en el núcleo del lemnisco lateral, pero muchas otras se lo saltan y viajan hasta el colículo inferior, donde todas, o casi todas, las fibras auditivas realizan sinapsis. A partir de allí, la vía sigue hacia el núcleo geniculado medial, donde todas las fibras hacen sinapsis. Finalmente, la vía continúa por medio de la radiación auditiva hasta la corteza auditiva, que ocupa básicamente la circunvolución superior del lóbulo temporal. Hay que reseñar varios aspectos importantes. En primer lugar, las señales procedentes de los dos oídos viajan por las vías de ambos lados del encéfalo, con un predominio de la transmisión a través de la vía contralateral. Como mínimo en tres lugares del tronco del encéfalo tiene lugar el cruce entre ambas vías: 1) en el cuerpo trapezoide; 2) en la comisura entre los dos núcleos del lemnisco lateral, y 3) en la comisura que conecta los dos colículos inferiores. En segundo lugar, muchas fibras colaterales de los fascículos auditivos pasan directamente al sistema reticular de activación en el tronco del encéfalo. Este sistema envía unas proyecciones difusas ascendentes por el tronco del encéfalo y descendentes hacia la médula espinal, y activa todo el sistema nervioso como respuesta a los sonidos fuertes. Otras colaterales van hacia el vermis del cerebelo, que también experimenta una activación instantánea en caso de un ruido brusco. En tercer lugar, los fascículos de fibras conservan un gran nivel de orientación espacial desde la cóclea a lo largo de todo el trayecto hasta la corteza. En realidad, existen tres representaciones espaciales de terminación para las diversas frecuencias sonoras en los núcleos cocleares, dos representaciones en los colículos inferiores, una representación precisa para las distintas frecuencias de sonido en la corteza auditiva, y un mínimo de otras cinco menos precisas en la corteza auditiva y las áreas auditivas de asociación.
Distinción de los «patrones» sonoros en la corteza auditiva La extirpación bilateral completa de la corteza auditiva no impide que un gato o un mono detecten los sonidos o generen una reacción no elaborada frente a su presencia. Sin embargo, reduce mucho o en ocasiones incluso llega a abolir la capacidad del animal para distinguir los diferentes tonos de sonido y sobre todo los patrones sonoros. Por ejemplo, un animal que haya recibido un entrenamiento para reconocer una combinación o una secuencia de tonos, uno detrás de otro según un patrón concreto, pierde esta capacidad cuando se destruye la corteza auditiva; además, el animal ya no puede volver a aprender este tipo de respuesta. Por tanto, la corteza auditiva posee una importancia especial para la distinción de los patrones de sonido tonales o secuenciales. La destrucción de las dos cortezas auditivas primarias en el ser humano reduce en gran medida la sensibilidad a la audición. Su desaparición en un solo lado únicamente disminuye un poco esta propiedad en el oído opuesto, pero no causa una sordera por las numerosas conexiones cruzadas que existen de un lado a otro en la vía nerviosa auditiva. Sin embargo, sí que afecta a la capacidad para localizar la fuente de un sonido, debido a que para cumplir esta función hacen falta las señales comparadas de ambas corteza. Determinación de la dirección de la que procede el sonido Una persona determina la dirección horizontal de la que viene el sonido por dos medios principales: 1) el lapso de tiempo transcurrido entre la llegada del sonido a un oído y al opuesto, y 2) la diferencia entre las intensidades de los sonidos en los dos oídos. El primer mecanismo funciona mejor a frecuencias por debajo de 3.000 ciclos/s, y el segundo a frecuencias más altas debido a que la cabeza constituye una barrera mayor para el sonido en esta gama. El mecanismo del intervalo de tiempo distingue la dirección con mucha mayor exactitud que el mecanismo de la intensidad porque no depende de factores ajenos sino solo del plazo temporal exacto que haya pasado entre las dos señales acústicas. Si una persona está mirando directamente hacia la fuente del sonido, este llega a los dos oídos justo en el mismo instante, mientras que si el oído derecho está más cerca que el izquierdo, las señales sonoras del primero penetran en el encéfalo antes que las del segundo.
Mecanismos nerviosos para detectar la dirección del sonido La destrucción de la corteza auditiva a ambos lados del cerebro, tanto en el ser humano como en los mamíferos inferiores, provoca una pérdida casi completa de la capacidad para detectar la dirección de la que procede el sonido. Con todo, los análisis nerviosos encargados de este proceso de detección comienzan en los núcleos olivares superiores del tronco del encéfalo, aunque hace falta la integridad de la vía nerviosa que va desde estos núcleos hasta la corteza para la interpretación de las señales. Se piensa que el mecanismo es el siguiente. El núcleo olivar superior se divide en dos componentes: 1) el núcleo olivar superior medial, y 2) el núcleo olivar superior lateral. El núcleo lateral se ocupa de detectar la dirección de la que viene el sonido, posiblemente mediante la simple comparación entre la diferencia de las intensidades sonoras que llegan a ambos oídos y el envío de la señal correspondiente hacia la corteza auditiva para calcular la dirección. Sin embargo, el núcleo olivar superior medial posee un mecanismo específico para detectar el lapso de tiempo transcurrido entre las señales acústicas que penetran por los dos oídos. Este núcleo contiene una gran cantidad de neuronas que presentan dos dendritas principales, una que proyecta hacia la derecha y la otra hacia la izquierda. La señal acústica procedente del oído derecho incide sobre la dendrita derecha, y la del oído izquierdo lo hace sobre la dendrita izquierda. La intensidad de la excitación de cada neurona presenta una gran sensibilidad al intervalo de tiempo específico transcurrido entre las dos señales acústicas de ambos oídos. Las neuronas cercanas a uno de los bordes del núcleo generan su respuesta máxima cuando el lapso de tiempo es corto, mientras que las que están situadas próximas al borde opuesto responden a un intervalo largo; las que quedan entre ambas reaccionan con plazos intermedios de tiempo. Por tanto, cuando surge un patrón espacial de estimulación neuronal en el núcleo olivar superior medial, en el que el sonido viene directamente desde un punto delante de la cabeza, provoca un estímulo máximo en una colección de neuronas olivares, y cuando llega formando diferentes ángulos laterales activa otros grupos de neuronas en lados opuestos. Esta orientación espacial de las señales se transmite a continuación hasta la corteza auditiva, donde la dirección del sonido se determina según el lugar ocupado por las neuronas que envían un estímulo máximo. Se cree que todas estas señales encargadas de identificar la dirección del sonido se transmiten a través de una vía diferente y excitan un punto distinto en la corteza cerebral que la vía de transmisión y el lugar de terminación dedicados a los patrones tonales del sonido.
Señales centrífugas desde el sistema nervioso central hasta los centros auditivos inferiores Se han descubierto unas vías retrógradas a todos los niveles del sistema nervioso auditivo desde la corteza hasta la cóclea en el oído. La vía final básicamente va desde el núcleo olivar superior hasta las células ciliadas receptoras del sonido en el órgano de Corti. Estas fibras retrógradas poseen un carácter inhibidor. En efecto, se ha demostrado que la estimulación directa de puntos aislados en el núcleo olivar inhibe zonas específicas del órgano de Corti, al reducir sus sensibilidades sonoras de 15 a 20 decibelios. No cuesta entender cómo este hecho puede permitir a una persona encaminar su atención hacia sonidos de una cualidad particular mientras rechaza los que posean otras cualidades. Esta característica queda de manifiesto con facilidad cuando la escucha dentro de una orquesta sinfónica se centra en un solo instrumento. Alteraciones de la audición Tipos de sordera La sordera suele dividirse en dos tipos: 1) la que está causada por una alteración de la cóclea o del nervio coclear, o de los circuitos del sistema nervioso central del oído, que suele clasificarse como «sordera nerviosa», y 2) la ocasionada por la afectación de las estructuras físicas del oído que conducen el propio sonido hasta la cóclea, lo que normalmente se denomina «sordera de conducción». Si se destruye la cóclea o el nervio coclear, la persona sufre una sordera permanente. Sin embargo, si ambas estructuras están aún íntegras pero ha desaparecido o se ha anquilosado el sistema tímpanohuesecillos (se ha «congelado» en su lugar por una fibrosis o una calcificación), las ondas sonoras aún pueden llegar hasta la cóclea por medio de la conducción ósea desde un generador del sonido aplicado sobre el cráneo encima del oído. Audímetro Para determinar la naturaleza de cualquier incapacidad auditiva se emplea el «audímetro». Este instrumento es un audífono conectado a un oscilador electrónico capaz de emitir tonos puros que abarquen desde las frecuencias más bajas hasta las más altas y se calibra de modo que el sonido con un nivel de intensidad nulo a cada frecuencia sea el volumen que apenas puede escucharse con un oído normal. Un mecanismo calibrado para controlar el volumen puede incrementarlo más allá del valor cero. Si el volumen ha de elevarse 30 decibelios por encima de lo normal antes de que sea posible escucharlo, se dice que la persona tiene una hipoacusia de 30 decibelios para esa frecuencia concreta. Al efectuar una prueba auditiva mediante un audímetro, se exploran unas 8 a 10 frecuencias que cubren todo el espectro audible, y se determina la pérdida de audición para cada una de ellas. De este modo se traza el denominado audiograma. Audiograma para la sordera de conducción en el oído medio Un tipo frecuente de sordera está causado por la fibrosis del oído medio después de haber sufrido infecciones repetidas o por la que acontece en la enfermedad hereditaria llamada otoesclerosis. En cualquier caso, las ondas sonoras no pueden transmitirse con facilidad a través de los huesecillos desde la membrana timpánica hasta la ventana oval.
El ojo: I. Óptica de la visión. PRINCIPIOS FISICOS DE LA OPTICA. Refracción de la luz Índice de refracción de un medio transparente Los rayos de luz viajan a través del aire a una velocidad de unos 300.000 km/s, pero se desplazan con mucha mayor lentitud cuando recorren sólidos y líquidos transparentes. El índice de refracción de una sustancia transparente es el cociente entre la velocidad de la luz en el aire y su velocidad en ese medio. El valor que toma en el aire es de 1. Por tanto, si la luz atraviesa un tipo concreto de vidrio a una velocidad de 200.000 km/s, el índice de refracción de este material es 300.000 dividido por 200.000, es decir, 1,5. Refracción de los rayos de luz en la superficie de transición entre dos medios con índices de refracción diferentes Cuando un rayo de luz que avance en un haz choca contra una superficie limitante que quede perpendicular a su llegada, penetra en el segundo medio sin desviarse de su trayectoria. Los únicos efectos que acontecen son un descenso de la velocidad de transmisión y una reducción de la longitud de onda, tal como se observa en la imagen por las distancias más cortas existentes entre los frentes de onda.
Cuando el rayo de luz atraviesa una superficie de separación que forma un ángulo cambia de dirección si los índices de refracción de ambos medios son diferentes entre sí. En esta imagen concreta, los rayos de luz abandonan el aire, cuyo índice de refracción es 1, y entran en un trozo de vidrio con un índice de refracción de 1,5. Cuando el haz choca por primera vez contra la superficie de contacto en ángulo, su borde inferior entra en el vidrio antes que el superior. El frente de onda de la porción alta del haz sigue viajando a una velocidad de 300.000 km/s, mientras que el que ya ha penetrado en el vidrio lo hace a 200.000 km/s. Esta diferencia en la velocidad implica que su porción superior se adelanta a la inferior, de manera que el frente deja de ser vertical y se angula hacia la derecha. Dado que la dirección con la que viaja la luz siempre es perpendicular al plano formado por el frente de onda, la trayectoria de avance del haz luminoso se inclina hacia abajo.
Una lente convexa concentra los rayos de luz La entrada a una lente convexa de unos rayos de luz paralelos. Los rayos luminosos que inciden sobre el centro de la lente chocan exactamente perpendiculares contra su superficie y, por tanto, la atraviesan sin sufrir ninguna refracción. Sin embargo, al alejarse hacia cualquiera de los bordes de la lente los rayos tropiezan con una superficie que forma un ángulo paulatinamente mayor. Los más externos se desvían cada vez más hacia el centro, lo que se denomina convergencia de los rayos. La mitad de su giro sucede al entrar en la lente y la otra mitad tiene lugar al salir por el lado opuesto. Si la lente tiene exactamente la curvatura adecuada, los rayos paralelos que atraviesan cada parte de la misma se desviarán justo lo suficiente para que todos se crucen en el mismo sitio, que se llama punto focal.
En el esquema central, los rayos luminosos que penetran en la lente convexa en vez de ser paralelos son divergentes debido a que el origen de la luz es una fuente puntual que no está lejos de la propia lente. Dado que estos rayos divergen a medida que se separan de su punto de origen, en el dibujo puede observarse que no se reúnen a la misma distancia de la lente que los rayos paralelos. Con otras palabras, cuando los rayos de luz que ya son divergentes llegan a una lente convexa, la distancia hasta el foco en el lado opuesto de la lente es más larga que la distancia focal de los rayos paralelos. El poder dióptrico de las lentes cóncavas no se puede establecer en función de la distancia focal existente después de atravesarla porque los rayos luminosos divergen en vez de concentrarse en un punto. Sin embargo, si dispersa los rayos de luz en la misma proporción en que una lente convexa de 1 dioptría los reúne, se dice que la lente cóncava tiene una potencia dióptrica de –1. Análogamente, si separa los rayos luminosos tanto como los concentra una lente de +10 dioptrías, se habla de que posee una potencia de –10 dioptrías. Las lentes cóncavas «neutralizan» el poder dióptrico de las convexas. Por tanto, si se coloca una lente cóncava de 1 dioptría justo delante de una lente convexa de 1 dioptría, esto crea un sistema de lentes con un poder dióptrico nulo. La potencia de las lentes cilíndricas se calcula de la misma manera que en el caso de las lentes esféricas, salvo por la necesidad de consignar el eje de las primeras además de su potencia. Por el contrario, si es de tipo cóncavo y causa una divergencia de los rayos luminosos equivalente a la convergencia ocasionada por otra lente cilíndrica de +1 dioptría, tiene una potencia de –1 dioptría. Si la línea enfocada es horizontal, se dice que su eje es de 0°. Si fuera vertical, su eje sería de 90°. Óptica del ojo «El ojo como una cámara «. El ojo equivale a una cámara fotográfica corriente desde el punto de vista óptico. Posee un sistema de lentes, un sistema de apertura variable (la pupila) y una retina que corresponde a la película. El sistema ocular de lentes está compuesto por cuatro superficies de refracción: 1) la separación entre el aire y la cara anterior de la córnea 2) la separación entre la cara posterior de la córnea y el humor acuoso 3) la separación entre el humor acuoso y la cara anterior del cristalino 4) la separación entre la cara posterior del cristalino y el humor vítreo. El índice de refracción para el aire es 1; el de la córnea, 1,38; el del humor acuoso, 1,33; el del cristalino (como promedio), 1,4, y el del humor vítreo, 1,34.
Consideración de todas las superficies oculares de refracción como una sola lente: «reducción» del ojo Si todas las superficies oculares de refracción se suman algebraicamente y a continuación se tratan como una sola lente, la óptica del ojo normal puede simplificarse y representarse de forma esquemática en una «reducción del ojo». Esta representación resulta práctica para realizar cálculos sencillos. En la reducción del ojo se considera que existe una sola superficie de refracción, con su punto central 17 mm por delante de la retina y un poder dióptrico total de 59 dioptrías cuando la acomodación del cristalino corresponde a la visión de lejos. La cara anterior de la córnea (y no el cristalino) aporta aproximadamente dos tercios de las 59 dioptrías del poder dióptrico ocular. La principal razón de este fenómeno estriba en que el índice de refracción de la córnea es sensiblemente distinto al del aire, mientras que el del cristalino no presenta grandes diferencias con los índices del humor acuoso y el humor vítreo. El poder dióptrico total del cristalino en el interior del ojo, bañado normalmente por líquido a ambos lados, solo es de 20 dioptrías, más o menos la tercera parte del poder dióptrico ocular total. Sin embargo, la importancia de este elemento radica en que, como respuesta a las señales nerviosas procedentes del encéfalo, su curvatura puede aumentar notablemente para permitir la «acomodación». Formación de una imagen en la retina De la misma manera que una lente de vidrio es capaz de enfocar una imagen sobre una hoja de papel, el sistema ocular de lentes puede enfocar una imagen sobre la retina. El resultado está dado la vuelta e invertido con respecto al objeto. Sin embargo, la mente percibe los objetos en su posición derecha a pesar de su orientación al revés en la retina debido a que el cerebro está entrenado para considerar como normal una imagen invertida. Mecanismo de «acomodación» En los niños, el poder dióptrico del cristalino puede aumentar a voluntad desde 20 dioptrías hasta unas 34, lo que corresponde a una «acomodación» de 14 dioptrías. Para conseguirlo, su forma cambia desde una lente con una convexidad moderada hasta una lente muy convexa. En una persona joven, el cristalino está compuesto por una potente cápsula elástica rellena de un líquido viscoso de carácter proteináceo, pero transparente. Cuando se encuentra en estado de relajación, sin ninguna tensión aplicada sobre la cápsula, adopta una forma casi esférica, debido básicamente a la retracción elástica de este elemento.
La estimulación de los nervios parasimpáticos contrae los dos tipos de fibras que componen el músculo ciliar, lo que relaja los ligamentos del cristalino y propicia un aumento del grosor y del poder dióptrico de dicha estructura. Con el incremento del poder dióptrico, el ojo enfoca objetos más cercanos que cuando posee un poder menor. Por consiguiente, a medida que se aproxima un objeto distante hacia el ojo, la cantidad total de impulsos parasimpáticos que inciden sobre el músculo ciliar ha de crecer de forma progresiva para mantener el objeto constantemente enfocado. (La estimulación simpática ejerce un efecto añadido para la relajación del músculo ciliar, pero esta acción resulta tan débil que casi no desempeña ninguna función en el mecanismo normal de la acomodación.
Presbicia: pérdida de acomodación en el cristalino A medida que una persona envejece, el cristalino crece y se engruesa perdiendo mucha elasticidad, en parte debido a la desnaturalización progresiva de sus proteínas. La capacidad que posee de modificar su forma disminuye con la edad. El poder de acomodación desciende de unas 14 dioptrías en un niño hasta menos de 2 para la época en que una persona llega a los 45 o 50 años y desciende prácticamente hasta 0 dioptrías con 70 años de edad.
Errores de refracción Emetropía (visión normal)
el ojo se considera normal, o «emétrope», si los rayos de luz paralelos procedentes de objetos alejados quedan enfocados con nitidez en la retina cuando el músculo ciliar esté relajado por completo. Esto significa que el ojo emétrope es capaz de ver todos los objetos distantes con claridad mientras el músculo ciliar se mantiene relajado. Sin embargo, para enfocar los objetos de cerca, el ojo ha de contraer el músculo ciliar y aportar así el grado de acomodación oportuno.
Hipermetropía La hipermetropía suele deberse a la presencia de un globo ocular demasiado corto o, a veces, de un sistema de lentes demasiado débil, el sistema de lentes relajado no desvía lo suficiente los rayos de luz paralelos como para que lleguen a estar enfocados en el momento de alcanzar la retina. Para vencer esta anomalía, el músculo ciliar ha de contraerse con el fin de incrementar la potencia del cristalino. Una persona hipermétrope, cuando recurre al mecanismo de la acomodación, es capaz de enfocar los objetos alejados en la retina. Si no ha consumido más que una pequeña parte de la potencia que ofrece el músculo ciliar para acomodar la vista a los objetos distantes, aún le queda un gran poder de acomodación, y también puede enfocar con nitidez otros objetos cada vez más cercanos al ojo hasta llegar al límite de contracción del músculo ciliar. En la vejez, cuando el cristalino haya adquirido una «presbicia», una persona hipermétrope normalmente es incapaz de acomodar lo suficiente para enfocar siquiera los objetos alejados, y mucho menos aún los cercanos. Miopía En la miopía, cuando el músculo ciliar está relajado del todo, los rayos de luz procedentes de objetos alejados quedan enfocados delante de la retina. Este trastorno suele deberse a la existencia de un globo ocular demasiado largo, pero puede obedecer a la acción de un poder dióptrico excesivo en el sistema ocular de lentes. No existe ningún mecanismo por el que el ojo sea capaz de reducir la potencia del cristalino por debajo de la que existe cuando el músculo ciliar se encuentra relajado por completo. Las personas miopes (o «cortas de vista») carecen de una herramienta para enfocar con nitidez los objetos alejados sobre la retina. Sin embargo, a medida que se acerca el objeto al ojo, acaba por aproximarse lo suficiente como para poder enfocar su imagen. Más tarde, si llega aún más cerca del ojo, puede apelarse al mecanismo de la acomodación para conservar la imagen enfocada con claridad. Una persona miope posee un «punto lejano» concreto que es el límite para la visión nítida. Corrección de la miopía y de la hipermetropía mediante el empleo de lentes Si las superficies oculares de refracción poseen demasiado poder dióptrico, como en la miopía, este exceso puede neutralizarse colocando una lente cóncava esférica delante del ojo, que provocará la divergencia de los rayos.
Hipermetropía La hipermetropía suele deberse a la presencia de un globo ocular demasiado corto o, a veces, de un sistema de lentes demasiado débil, el sistema de lentes relajado no desvía lo suficiente los rayos de luz paralelos como para que lleguen a estar enfocados en el momento de alcanzar la retina. Para vencer esta anomalía, el músculo ciliar ha de contraerse con el fin de incrementar la potencia del cristalino. Una persona hipermétrope, cuando recurre al mecanismo de la acomodación, es capaz de enfocar los objetos alejados en la retina. Si no ha consumido más que una pequeña parte de la potencia que ofrece el músculo ciliar para acomodar la vista a los objetos distantes, aún le queda un gran poder de acomodación, y también puede enfocar con nitidez otros objetos cada vez más cercanos al ojo hasta llegar al límite de contracción del músculo ciliar. En la vejez, cuando el cristalino haya adquirido una «presbicia», una persona hipermétrope normalmente es incapaz de acomodar lo suficiente para enfocar siquiera los objetos alejados, y mucho menos aún los cercanos. Miopía En la miopía, cuando el músculo ciliar está relajado del todo, los rayos de luz procedentes de objetos alejados quedan enfocados delante de la retina. Este trastorno suele deberse a la existencia de un globo ocular demasiado largo, pero puede obedecer a la acción de un poder dióptrico excesivo en el sistema ocular de lentes. No existe ningún mecanismo por el que el ojo sea capaz de reducir la potencia del cristalino por debajo de la que existe cuando el músculo ciliar se encuentra relajado por completo. Las personas miopes (o «cortas de vista») carecen de una herramienta para enfocar con nitidez los objetos alejados sobre la retina. Sin embargo, a medida que se acerca el objeto al ojo, acaba por aproximarse lo suficiente como para poder enfocar su imagen. Más tarde, si llega aún más cerca del ojo, puede apelarse al mecanismo de la acomodación para conservar la imagen enfocada con claridad. Una persona miope posee un «punto lejano» concreto que es el límite para la visión nítida. Corrección de la miopía y de la hipermetropía mediante el empleo de lentes Si las superficies oculares de refracción poseen demasiado poder dióptrico, como en la miopía, este exceso puede neutralizarse colocando una lente cóncava esférica delante del ojo, que provocará la divergencia de los rayos.
Astigmatismo El astigmatismo es un error de la refracción ocular que hace que la imagen visual de un plano quede enfocada a una distancia diferente de la que corresponde a su plano perpendicular. Lo más frecuente es que el astigmatismo obedezca a una curvatura de la córnea demasiado grande en uno de los planos del ojo. Un ejemplo de lente astigmática sería una superficie como la de un huevo situado de lado frente a la luz incidente. El grado de curvatura que presenta el plano correspondiente al eje mayor del huevo no llega a ser tan acusado como el del correspondiente al eje menor. El ojo astigmático puede plantearse como si su sistema de lentes estuviera constituido por dos lentes cilíndricas de potencia diferente colocadas perpendiculares entre sí. Para corregir esta situación, el procedimiento habitual consiste en encontrar por ensayo y error una lente esférica que sea capaz de corregir el foco en uno de los dos planos de la lente astigmática. Entonces, para corregir el error restante en el plano que queda se recurre a una lente cilíndrica más. Con este fin, hay que determinar el eje y la potencia de la lente cilíndrica requerida
Corrección de las anomalías ópticas con lentillas Las lentes de contacto de cristal o de plástico pueden colocarse hasta acoplarlas perfectamente encajadas contra la cara anterior de la córnea. Estas lentes se mantienen en su sitio mediante una fina película de líquido lagrimal que llene el espacio entre la lente de contacto y la superficie anterior del ojo. Un rasgo especial de la lente de contacto es que anula casi por completo la refracción que se produce normalmente en la cara anterior de la córnea. La razón de esta anulación estriba en que las lágrimas situadas entre la lente de contacto y la córnea tienen un índice de refracción prácticamente igual que el de esta última, por lo que su cara anterior deja de ocupar un lugar de relieve en el sistema óptico del ojo. En su lugar, es la cara externa de la lentilla la que desempeña la función principal. Por tanto, la refracción de esta superficie sustituye a la habitual de la córnea. Este factor resulta especialmente importante en las personas cuyos errores de refracción oculares están ocasionados por una forma anormal de esta estructura, como los que tienen una córnea abombada de configuración extraña, proceso denominado queratocono. Sin la lente de contacto, la disposición saliente de la córnea origina una alteración tan acusada de la visión que casi es imposible corregirla satisfactoriamente con gafas; sin embargo, cuando se recurre a una lentilla, se neutraliza la refracción de la córnea, que queda reemplazada por la refracción normal de la nueva superficie externa. La lente de contacto ofrece varias ventajas más, entre ellas el hecho de que: 1) gira con el ojo y aporta un campo de visión nítida más amplio que las gafas, y 2) ejerce escasos efectos sobre las dimensiones del objeto observado por la persona a su través, mientras que las lentes colocadas en torno a 1 cm delante del ojo influyen sobre el tamaño de la imagen, además de corregir el foco. Cataratas: zonas opacas en el cristalino Las «cataratas» son una alteración ocular especialmente frecuente que sucede sobre todo en las personas mayores. Una catarata consiste en una o varias zonas turbias u opacas en el interior del cristalino. Durante la primera etapa de su formación se desnaturalizan las proteínas de algunas fibras del cristalino. Más adelante, estas mismas proteínas se coagulan para generar áreas opacas en lugar de las fibras proteicas transparentes normales. Cuando una catarata ha oscurecido la transmisión de luz tanto como para afectar seriamente la visión, la situación puede corregirse mediante la extirpación quirúrgica del cristalino. Si se lleva a cabo esta intervención, el ojo pierde una gran parte de su poder dióptrico, que ha de restituirse mediante la colocación de una lente convexa potente delante del mismo; sin embargo, por lo común se implanta una lente artificial de plástico dentro del ojo en el lugar que ocupaba el cristalino extraído. Agudeza visual En teoría, la luz procedente de una fuente puntual alejada, cuando se enfoca sobre la retina, debería ser infinitamente pequeña. Sin embargo, dado que el sistema ocular de lentes nunca es perfecto, dicho punto retiniano corrientemente posee un diámetro total de unos 11 μm, incluso cuando el ojo presenta un sistema óptico normal dotado de su máxima resolución. La mancha resulta más brillante en su centro y se va oscureciendo gradualmente hacia los bordes. El diámetro medio de los conos en la fóvea de la retina, que es su porción central, donde la visión está más desarrollada, es de unos 1,5 μm, lo que supone la séptima parte del diámetro del punto luminoso. No obstante, como la mancha de luz tiene un núcleo central brillante y se difumina hacia los bordes, una persona normalmente puede distinguir dos puntos separados si su centro queda a un mínimo de 2 μm de distancia en la retina, lo que excede ligeramente la anchura de los conos en la fóvea. Procedimiento clínico para establecer la agudeza visual La tabla de exploración ocular suele constar de letras de diferentes tamaños colocadas a 6 m de la persona examinada. Se dice que la visión de esta persona es de 20/20 (una visión normal) si puede ver bien las letras de unas dimensiones que debería ser capaz de ver a esa distancia. Si esto no sucede hasta mostrarle letras que debería ser capaz de ver a 60 m, se dice que tiene una visión de 20/200. Dicho de otro modo, el procedimiento clínico para expresar la agudeza visual consiste en utilizar una fracción matemática que contenga el cociente entre dos distancias, lo que también corresponde al cociente entre la agudeza visual de un individuo y la de una persona con una agudeza normal.
Determinación de la distancia de un objeto al ojo: «percepción de la profundidad» Una persona normalmente percibe la distancia por tres medios principales: 1) el tamaño que poseen las imágenes de los objetos conocidos sobre la retina; 2) el efecto del movimiento de paralaje, y 3) el fenómeno de la estereopsia. Esta capacidad para determinar la distancia se llama percepción de la profundidad. Determinación de la distancia según el tamaño de las imágenes retinianas de objetos conocidos Si se sabe que una persona observada mide 1,8 m de altura, puede determinarse la distancia a la que se encuentra simplemente por el tamaño de su imagen en la retina. Uno no piensa conscientemente en ello, pero el cerebro ha aprendido a calcular automáticamente la distancia de los objetos según las dimensiones de una imagen cuando se conocen sus medidas. Determinación de la distancia mediante el movimiento de paralaje Otro medio importante al que recurren los ojos para determinar la distancia es el movimiento de paralaje. Si una persona mira hacia lo lejos con los ojos completamente quietos, no percibe este fenómeno, pero cuando desplaza su cabeza hacia un lado o hacia el otro, las imágenes de los objetos cercanos se mueven con rapidez a través de la retina, mientras que en el caso de los objetos alejados permanecen casi inmóviles del todo. Por ejemplo, al apartar la cabeza 3 cm hacia un lado cuando un objeto se encuentra solo a 3 cm delante del ojo, su imagen recorre casi toda la retina, mientras que la de otro objeto a 60 m de distancia no muestra ningún desplazamiento perceptible. Por tanto, mediante este mecanismo del movimiento de paralaje, puede afirmarse la distancia relativa de los diferentes objetos, aunque no se esté utilizando más que un ojo. Determinación de la distancia por estereopsia: visión binocular Otro método por el que se percibe el paralaje es el de la «visión binocular». Dado que un ojo está a poco más de 5 cm del otro, las imágenes formadas en las dos retinas son diferentes entre sí. Por ejemplo, un objeto a 3 cm delante de la nariz crea una imagen en el lado izquierdo de la retina del ojo izquierdo y en el lado derecho de la retina del ojo derecho, mientras que la imagen de otro objeto pequeño 6 m por delante de la nariz ocupa puntos prácticamente correspondientes en el centro de las dos retinas.
Oftalmoscopio El oftalmoscopio es un instrumento por el que un observador puede mirar dentro del ojo de otra persona y ver la retina con claridad.
Si en la retina de un ojo emétrope hay un punto de luz brillante, los rayos luminosos divergen desde él hacia el sistema ocular de lentes. Después de atravesarlo, son paralelos entre sí porque la retina está separada una distancia focal por detrás de dicho sistema. A continuación, cuando estos rayos paralelos entran en el ojo emétrope de otra persona, se concentran de nuevo en un foco puntual de la retina de esta segunda persona, porque en ella también queda a una distancia focal por detrás de la lente. Cualquier punto de luz en la retina del ojo observado se proyecta a un punto focal en la retina del ojo observador. Por tanto, si se hace que la retina de una persona emita luz, su imagen quedará enfocada en la retina del observador, siempre que los dos ojos sean emétropes, nada más que con que estén mirándose el uno al otro. Si el poder dióptrico del ojo observado o del ojo del observador no es normal, resulta necesario corregirlo para que el observador vea una imagen nítida de la retina examinada. Un oftalmoscopio normal posee una serie de lentes muy pequeñas montadas en un revólver de manera que pueda pasarse de una a otra hasta que se logre corregir la refracción anormal mediante la selección de una lente con la potencia adecuada. En una persona joven normal, los reflejos naturales de acomodación provocan un aumento aproximado de +2 dioptrías en la potencia del sistema de lentes de cada ojo. Para subsanar esta situación es necesario rotar el revólver de las lentes aproximadamente hasta una corrección de –4 dioptrías. Salida del humor acuoso desde el ojo Una vez que se ha formado el humor acuoso en los procesos ciliares, primero fluye a través de la pupila hacia la cámara anterior del ojo. Tiene una membrana endotelial tan porosa que incluso las moléculas grandes de proteínas pueden pasar desde la cámara anterior hacia el conducto de Schlemm, lo mismo que las pequeñas partículas materiales con un tamaño hasta el de los eritrocitos. Aunque en realidad se trate de un vaso sanguíneo venoso, normalmente es tan grande la cantidad de humor acuoso que entra en el conducto de Schlemm que este líquido lo llena por completo en vez de la sangre. Los pequeños vasos que se extienden desde el conducto de Schlemm hasta las venas más grandes del ojo solo suelen contener humor acuoso, y se denominan venas acuosas. Presión intraocular La presión intraocular normal media es de unos 15 mmHg, con un intervalo desde 12 hasta 20 mmHg. El glaucoma es un trastorno en el que la presión intraocu- lar puede alcanzar cifras peligrosamente elevadas (del orden de 60 a 70 mmHg). A medida que la presión asciende por encima de 20 a 30 mmHg, los axones de las células ganglionares de la retina que forman el nervio óptico se comprimen hasta el extremo de interrumpir el flujo axónico, causando un daño permanente a una neurona originaria.
El ojo: II. Función receptora y nerviosade la retina.
La retina se compone de 10 capas o estratos celulares quese enumeran de forma secuencial, comenzando por el másexterno (el más alejado del centro del globo ocular): 1. Capa pigmentaria 2. Capa de conos y bastones. 3. Membrana limitante externa. 4. Capa nuclear externa 5. Capa plexiforme externa 6. Capa nuclear interna. 7. Capa plexiforme interna 8. Capa ganglionar 9. Capa de fibras del nervio óptico. 10. Membrana limitante interna. Cuando la luz atraviesa el sistema ocular de lentes,encuentra primero la membrana limitante interna, las fibras del nervio óptico y la capa de células ganglionares; luego prosigue por las demás capas hasta alcanzar los receptores(conos y bastones). La fóvea es una región especializada, de aproximadamente 1 mm2, situada en el centro de la retina.Dentro del centro de la fóvea existe una zona, de nominada fóvea central, con un diámetro de 0,3 mm; esta es la región donde la agudeza visual alcanza el máximo y la capa de fotorreceptores solo contiene conos. Además, las capas subyacentes de la retina a lo largo de las fibras del nervio óptico y de los vasos sanguíneos se desplazan lateralmente para facilitar el acceso más directo posible a los receptores.
Cada fotorreceptor se compone de: 1) un segmentoexterno 2) un segmento interno 3) una región nuclear 4) el cuerpo o terminal sináptico. Los receptores se denominan bastones o conos, dependiendo, principalmente, de la forma del segmento externo.
El fotopigmento sensible a la luz rodopsina se localiza en elsegmento externo del bastón, mientras que el del cono con-tiene un material parecido, un pigmento sensible a los colores, la fotopsina. Estos fotopigmentos son proteínas incorporadas aun conjunto apilado de discos membranosos en el segmentoexterno del receptor, que representa un repliegue de la membrana celular externa. Sin embargo, no resulta fácil de reconocer en la porción distal del segmento externo del bastón,donde los discos membranosos se desprenden de manera secundaria y están totalmente contenidos dentro de la membrana limitante de ese segmento. Los segmentos internos de los bastones y conos son básicamente idénticos y contienen componentes y orgánulos citoplásmicos comunes a otros somas neuronales. Cada núcleo del fotorreceptor se continúa con su propio segmento interno, pero la membrana limitante externa de la retina establece una separación o frontera incompleta entre la capa de los segmentos internos y la capa de los núcleos fotorreceptores (capa nuclear externa). El cuerpo sináptico contiene elementos como mitocondrias y vesículas sinápticas, habituales en las terminaciones axónicas del cerebro. El pigmento negro melanina de la capa de pigmento reduce la reflexión de la luz por el globo ocular y cumple, por tanto, una función parecida al color negro del interior de los fuelles de una cámara. La importancia de este pigmento se reconoce mejor por su ausencia en las personas albinas. Los albinos casi nunca alcanzan una agudeza visual mayor de 20/100 debido a la enorme reflexión que tiene lugar dentro del globo ocular. La capa de pigmento también almacena grandes cantidades de vitamina A, empleadas en la síntesis de los pigmentos visuales. La arteria central de la retina irriga solo las capas más internas (desde los axones de las células ganglionares hasta la capa nuclear interna). Las capas más externas de la retina son perfundidas mediante difusión desde la coroides,tejido muy vascularizado que se sitúa entre la esclerótica y la retina.
FOTOQUIMICA DE LA VISION.
Ciclo rodopsina-retinal y excitación de los bastones. La rodopsina se descompone por la energía lumínica. El fotopigmento rodopsina de los bastones se concentra en la porción del segmento externo que sobresale hacia la capa pigmentaria. Esta sustancia es una combinación de la proteína escotopsina y del pigmento carotenoide retinal o, más concretamente,11-cis retinal. Cuando la rodopsina absorbe la energía lumínica,el retinal se transforma en su configuración todo-trans y comienzan a separarse los componentes retinal y escotopsina. Tras una serie de reacciones extraordinariamente rápidas, el componente retinal se transforma en lumirrodopsina, metarrodopsina I, metarrodopsina II y, por último, escotopsina; y se escinde el todo-trans retinal. Durante este proceso, se cree que la metarrodopsina II se ocupa de la transmisión de losimpulsos sucesivos por la retina. Regeneración de la rodopsina. En la primera etapa de regeneración de la rodopsina, el todo-trans retinal se convierte en 11-cis retinal, que se combina en seguida con la escotopsina para dar la rodopsina. Existe una segunda vía deformación de la rodopsina basada en la conversión del todo-trans retinal en todo-trans retinol, una forma de vitamina A. El retinol se convierte por vía enzimática en 11-cis retinol y luegoen 11-cis retinal, que puede combinarse con la escotopsinapara dar rodopsina. Si la retina contiene un exceso de retinal,se transforma en vitamina A, disminuyendo así la cantidad total de rodopsina de la retina. Las personas con carencia de vitamina A experimentan ceguera nocturna porque los bastones son los fotorreceptores que más se utilizan en condiciones de iluminación crepuscular y la formación de rodopsina disminuye de forma espectacular por la ausencia de vitamina A. Este trastorno se revierte en 1 h o menos mediante la inyección intravenosa de vitamina A.
Fotoquímica de la visión de los colores por los conos Anteriormente se ha señalado que las sustancias fotosensibles de los conos poseen casi exactamente la misma composición química que la rodopsina de los bastones. La única diferencia radica en que sus porciones proteicas u opsinas (llamadas fotopsinas en los conos) son un poco distintas de la escotopsina de los bastones. La porción de todos los pigmentos visuales correspondiente al retinal es absolutamente idéntica en los conos y en los bastones. Por tanto, los pigmentos sensibles al color de los conos consisten en combinaciones de retinal y fotopsinas. En la explicación sobre la visión de los colores que se ofrece más adelante en este capítulo, quedará patente que solo uno de los tres tipos de pigmentos destinados a este fin está presente en cualquiera de los diferentes conos, lo que les concede una sensibilidad selectiva a los diversos colores: azul, verde o rojo. Estos pigmentos para el color se llaman, respectivamente, pigmento sensible al azul, pigmento sensible al verde y pigmento sensible al rojo. Sus características de absorción en los tres tipos de conos muestran unas absorbencias máximas ante la luz de las longitudes de onda de 445, 535 y 570 nm, respectivamente. Regulación automática de la sensibilidad retiniana: adaptación a la luz y a la oscuridad Si una persona ha estado expuesta a una luz radiante durante horas, una gran parte de las sustancias fotosensibles en los conos y los bastones habrá quedado reducida a retinal y opsinas. Por añadidura, gran parte del retinal presente en ambos tipos de receptores se habrá convertido en vitamina A. Debido a estos dos efectos, las concentraciones de productos fotosensibles que quedan en los conos y los bastones son considerablemente bajas, y la sensibilidad del ojo a la luz está reducida de forma proporcional. Este proceso se llama adaptación a la luz. Por el contrario, si una persona permanece en la oscuridad durante un período prolongado, el retinal y las opsinas de los conos y los bastones han vuelto a convertirse en los pigmentos sensibles a la luz.
Valores de la adaptación visual a la luz y a la oscuridad Entre los límites de la adaptación máxima a la oscuridad y a la luz, el ojo puede modificar su sensibilidad lumínica hasta de 500.000 a 1 millón de veces, con una corrección automática ante los cambios de iluminación. Dado que la recepción de las imágenes por la retina requiere la detección de puntos luminosos y oscuros en la figura, resulta fundamental que su sensibilidad esté siempre ajustada de modo que los receptores respondan a las zonas más luminosas, pero no a las más oscuras. Un ejemplo de mala regulación de la adaptación retiniana que puede citarse ocurre cuando una persona sale de un cine y recibe la luz radiante del sol. En ese instante, incluso los puntos oscuros de las imágenes parecen extraordinariamente brillantes y, como consecuencia, la imagen visual en su integridad está decolorada, con un contraste escaso entre sus diferentes partes. Esta visión deficiente se prolonga hasta que la retina se haya adaptado lo suficiente para que las áreas más oscuras de la imagen dejen de estimular en exceso a los receptores.
Percepción de la luz blanca Una estimulación aproximadamente equivalente de los conos rojo, verde y azul da lugar a la sensación visual del blanco. Con todo, la luz no tiene una sola longitud de onda que corresponda al blanco; en su lugar, este color es una combinación de todas las longitudes del espectro. Además, la percepción del blanco puede lograrse mediante la estimulación de la retina con la combinación oportuna tan solo de tres colores escogidos que activen los tipos de conos respectivos más o menos por igual. Daltonismo Daltonismo rojo-verde El daltonismo rojo-verde es un trastorno genético que aparece casi exclusivamente en los varones. Es decir, los genes del cromosoma X femenino codifican para los conos respectivos. Con todo, el daltonismo casi nunca ocurre en las mujeres porque uno de los dos cromosomas X casi siempre posee como mínimo un gen normal para cada tipo de cono. Dado que el varón solo tiene un cromosoma X, la ausencia de un gen puede desembocar en un daltonismo. Como el cromosoma X del varón siempre se hereda de la madre, nunca del padre, esta alteración se transmite de la madre al hijo, y se dice que la madre es una portadora del daltonismo; aproximadamente el 8% de las mujeres son portadoras de daltonismo. Debilidad para el azul Solo rara vez faltan los conos azules, aunque en ocasiones están infrarrepresentados, lo que constituye un trastorno genéticamente hereditario que da origen al fenómeno llamado debilidad para el azul.
Función nerviosa de la retina Los diversos tipos neuronales son los siguientes: 1. Los fotorreceptores, los conos y los bastones, que transmiten las señales hacia la capa plexiforme externa, donde hacen sinapsis con las células bipolares y horizontales. 2. Las células horizontales, que transmiten las señales en sentido horizontal por la capa plexiforme externa desde los conos y los bastones hasta las células bipolares. 3. Las células bipolares, que transmiten las señales en sentido vertical desde los conos, los bastones y las células horizontales hacia la capa plexiforme interna, donde hacen sinapsis con las células ganglionares y amacrinas. 4. Las células amacrinas, que transmiten las señales en dos direcciones, directamente desde las células bipolares hasta las células ganglionares, u horizontalmente en el seno de la capa plexiforme interna desde los axones de las células bipolares hasta las dendritas de las células ganglionares o hasta otras células amacrinas. 5. Las células ganglionares, que transmiten las señales de salida desde la retina hacia el cerebro a través del nervio óptico.
La transmisión de la mayoría de los impulsos en las neuronas de la retina se produce por conducción electrotónica, no por potenciales de acción Las únicas neuronas de la retina que siempre transmiten señales visuales por medio de potenciales de acción son las células ganglionares, que las envían hasta el cerebro a través del nervio óptico. A veces, también se han registrado potenciales de acción en las células amacrinas, aunque su importancia sea dudosa. Por lo demás, todas las neuronas de la retina envían su información visual mediante conducción electrotónica, que puede explicarse del modo siguiente: la conducción electrotónica significa el flujo directo de una corriente eléctrica, y no de unos potenciales de acción, a lo largo del citoplasma neuronal y los axones nerviosos desde el punto de excitación hasta las sinapsis de salida. Incluso en los conos y los bastones, la conducción desde sus segmentos externos, donde se generan las señales visuales, hasta los cuerpos sinápticos es de tipo electrotónico. Es decir, cuando se produce la hiperpolarización como respuesta a la luz en el segmento externo de un cono o de un bastón, se transmite casi con la misma magnitud mediante un flujo de corriente eléctrica directo por el citoplasma hasta el cuerpo sináptico, sin que haga falta ningún potencial de acción. Entonces, cuando el transmisor de un cono o de un bastón estimula una célula bipolar u horizontal, una vez más la señal avanza desde la entrada hasta la salida por un flujo directo de corriente eléctrica, y no por potenciales de acción. La importancia de la transmisión electrotónica radica en que permite una conducción escalonada de la potencia de la señal. Por tanto, en el caso de los conos y los bastones, la magnitud del impulso de salida hiperpolarizante está directamente relacionada con la intensidad de la iluminación; no queda reducida a todo o nada, como sucedería con cada potencial de acción aislado.
Células bipolares despolarizantes e hiperpolarizantes Dos son los tipos de células bipolares que suministran señales excitadoras e inhibidoras opuestas en la vía visual: 1) la célula bipolar despolarizante, y 2) la célula bipolar hiperpolarizante. Es decir, algunas células bipolares se despolarizan con la excitación de los conos y los bastones, y otras se hiperpolarizan. Existen dos posibles explicaciones para esta diferencia. La primera señala que estas células bipolares pertenecen a dos clases totalmente distintas, de manera que una responde con una despolarización frente a la liberación del neurotransmisor glutamato por parte de los conos y los bastones, y la otra lo hace con una hiperpolarización. La opción alternativa defiende que una de las células bipolares recibe una excitación directa procedente de los conos y los bastones, mientras que el impulso llega a la otra por un camino indirecto a través de una célula horizontal. Dado que esta última es una célula inhibidora, se invertiría la polaridad de la respuesta eléctrica. Sea cual sea el mecanismo que siguen los dos tipos de respuestas, la importancia de este fenómeno reside en que permite que la mitad de las células bipolares envíe señales positivas y la otra mitad las envíe negativas.
Células amacrinas y sus funciones Se han identificado unas 30 clases de células amacrinas por medios morfológicos e histoquímicos. Ya se ha podido caracterizar la función más o menos de una docena de ellas y todas son diferentes. • Un tipo de célula amacrina forma parte de la vía directa para la visión de los bastones, a saber, la compuesta por el bastón, las células bipolares, las células amacrinas y las células ganglionares. • Otro tipo de célula amacrina responde potentemente cuando comienza una señal visual continua, pero su actividad se extingue con rapidez. • Otras células amacrinas presentan una respuesta enérgica al desaparecer las señales visuales, pero, una vez más, su descarga cesa a gran velocidad. • Otro tipo responde cuando se enciende o se apaga una luz, para indicar simplemente un cambio de iluminación, sin tener en cuenta su sentido. • Otro tipo más responde al movimiento de un punto a través de la retina en una dirección específica; por tanto, se dice que estas células amacrinas son sensibles a la dirección. Así pues, y en un cierto sentido, muchas de las células amacrinas o su mayoría son interneuronas que sirven para analizar las señales visuales antes de que lleguen a abandonar la retina.
Células ganglionares de la retina y sus campos respectivos Células W, X e Y Los estudios iniciales realizados en gatos describieron tres tipos distintos de células ganglionares de la retina, que se designan como células W, X e Y, basándose en sus diferencias en estructura y en función. Las células W envían señales por sus fibras en el nervio óptico a una velocidad lenta y reciben el mayor componente de su excitación desde los bastones, transmitida a través de pequeñas células bipolares y células amacrinas. Tienen campos amplios en la retina periférica, son sensibles para la detección del movimiento direccional en el campo de visión y, probablemente tienen importancia para la visión de los bastones en condiciones de oscuridad. Las células X poseen campos pequeños porque sus dendritas no ocupan una gran extensión en la retina, y así sus señales representan lugares separados de la retina y transmiten los detalles finos de las imágenes visuales. Asimismo, como cada célula X recibe conexiones al menos desde un cono, su actividad probablemente es responsable de la visión de los colores. Las células Y son las más grandes de todas y envían impulsos hacia el cerebro a 50 m/s o más rápidamente. Dado que poseen amplios campos dendríticos, las señales son captadas por estas células a partir de extensas zonas retinianas. Las células Y responden a las modificaciones rápidas de las imágenes visuales y comunican al sistema nervioso central la irrupción de un fenómeno nuevo en el campo visual de un modo casi instantáneo, pero no especifican con gran precisión su lugar de aparición, aparte de ofrecer indicios para que los ojos se desplacen hacia el estímulo excitador. Células P y M En los primates se utiliza una clasificación diferente de células ganglionares de la retina, y se han descrito hasta 20 tipos de estas células, cada uno de los cuales responde a una característica diferente de la escena visual. Algunas células responden mejor a direcciones específicas de movimiento u orientaciones, mientras que otras lo hacen a detalles finos, aumento o disminución de la luz o determinados colores. Las dos clases generales de células ganglionares de la retina más ampliamente estudiadas en los primates, incluidos los seres humanos, se designan como células magnocelulares (M) y parvocelulares (P). Las células P (también conocidas como células β o, en la retina central, como células ganglionares enanas) se proyectan hacia la capa de células parvocelulares (pequeñas) del núcleo geniculado lateral del tálamo. Las células M (conocidas también como células α o parasol) se proyectan a la capa magnocelular (células grandes) del núcleo geniculado lateral, que, a su vez, retransmite la información desde la cintilla óptica a la corteza visual, como se expone en el capítulo 52. Las diferencias principales entre las células P y M son las siguientes: 1. Los campos receptores de células P son mucho más pequeños que los de las células M. 2. Los axones de las células P conducen impulsos con mucha mayor lentitud que las células M.
Las respuestas de las células P a los estímulos, especialmente a los estímulos de colores, pueden ser sostenidas, mientras que las respuestas de las células M son mucho más transitorias. 4. Las células P son, en general, sensibles al color de un estímulo, mientras que las células M no lo son. 5. Las células M son mucho más sensibles que las P a estímulos en blanco y negro de bajo contraste. Las funciones principales de las células M y P son evidentes a partir de sus diferencias: las células P son muy sensibles a señales visuales que se relacionan con detalles finos y a diferentes colores, pero relativamente insensibles a señales de bajo contraste, mientras que las células M son muy sensibles a los estímulos de bajo contraste y a señales visuales en rápido movimiento. Se ha descrito un tercer tipo de célula ganglionar de la retina fotosensible que contiene su propio fotopigmento, la melanopsina. Se sabe mucho menos de este tipo celular, aunque estas células parecen enviar señales principalmente a zonas no visuales del encéfalo, en concreto el núcleo supraquiasmático del hipotálamo, el principal marcapasos circadiano. Supuestamente, estas señales ayudan a controlar el ritmo circadiano que sincroniza los cambios fisiológicos con la noche y el día. Excitación de las células ganglionares Potenciales de acción continuos y espontáneos en las células ganglionares Las células ganglionares son el punto de origen de las fibras largas que llegan al cerebro formando el nervio óptico. Dada la distancia que han de recorrer, el método de conducción electrotónico empleado por los conos, los bastones y las células bipolares en el interior de la retina deja de ser rentable; por tanto, las células ganglionares pasan a transmitir sus impulsos mediante potenciales de acción repetidos. Además, incluso cuando no están estimuladas, envían impulsos continuos a una frecuencia que oscila entre 5 y 40 por segundo. A su vez, las señales visuales quedan superpuestas sobre esta actividad de fondo de las células ganglionares.
Transmisión de las señales de color por parte de las células ganglionares Una sola célula ganglionar puede ser estimulada por varios conos o únicamente por unos pocos. Cuando los tres tipos de conos, rojo, azul y verde, activan la misma célula ganglionar, la señal transmitida por ella es idéntica ante cualquier color del espectro. Por tanto, este elemento no cumple ninguna función en la detección de los diversos colores. Más bien, se trata de una señal «blanca». En cambio, algunas células ganglionares reciben la excitación de un tipo de cono de un solo color, pero también la inhibición de un segundo tipo. Por ejemplo, este mecanismo sucede a menudo para los conos rojo y verde: el rojo provoca la excitación y el verde la inhibición o viceversa. Esta misma clase de efecto recíproco ocurre entre los conos azules por un lado y una combinación de los conos rojos y verdes (que se excitan con el amarillo) por el otro, lo que otorga una relación recíproca de excitación-inhibición entre los colores azul y amarillo. El mecanismo de este efecto opuesto producido por los colores es el siguiente: un tipo de cono de color excita la célula ganglionar por la vía excitadora directa a través de una célula bipolar despolarizante, mientras que el otro tipo de color la inhibe a través de la vía inhibidora indirecta mediante una célula bipolar hiperpolarizante. La importancia de estos mecanismos de contraste del color estriba en que representan un medio por el que la retina comienza a distinguir los colores. Así pues, cada tipo de célula ganglionar para el contraste de colores queda excitado por un color pero inhibido por el color «contrario». Por tanto, el análisis del color empieza en la retina y no supone una función íntegra del cerebro.
El ojo: III. Neurofisiología central de la visión. Vías visuales. Las señales nerviosas de este carácter abandonan la retina a través de los nervios ópticos. En el quiasma óptico, las fibras procedentes de la mitad nasal de la retina cruzan hacia el lado opuesto, donde se unen a las fibras originadas en la retina temporal contraria para formar los tractos ópticos o cintillas ópticas. A continuación, las fibras de cada tracto óptico hacen sinapsis en el núcleo geniculado lateral dorsal del tálamo, y desde allí, las fibras geniculocalcarinas se dirigen a través de la radiación óptica (también denominada tracto geniculocalcarino) hacia la corteza visual primaria en el área correspondiente a la cisura calcarina del lóbulo occipital medial.
Las fibras visuales también se dirigen a otras regiones más antiguas del encéfalo: 1) desde los tractos ópticos llega hasta el núcleo supraquiasmático del hipotálamo, se supone que para regular los ritmos circadianos que sincronizan los diversos cambios fisiológicos del organismo según la noche y
el día; 2) hacia los núcleos pretectales en el mesencéfalo, para suscitar movimientos reflejos de los ojos a fin de enfocarlos sobre los objetos de importancia y activar el reflejo fotomotor pupilar; 3) hacia el colículo superior, para controlar los movimientos direccionales rápidos de ambos ojos, y 4) hacia el núcleo geniculado lateral ventral del tálamo y las regiones basales adyacentes del cerebro, se cree que para contribuir al dominio de algunas de las funciones conductuales que lleva a cabo el organismo. Así pues, las vías visuales pueden dividirse a grandes rasgos en un sistema antiguo dirigido hacia el mesencéfalo y la base del prosencéfalo y un sistema nuevo para la transmisión directa de las señales visuales hacia la corteza visual situada en los lóbulos occipitales. En el ser humano, el sistema nuevo es el responsable de percibir prácticamente todos los aspectos de la forma visual, los colores y el resto de la visión consciente. A la inversa, en muchos animales primitivos, incluso la forma visual se detecta con el sistema más antiguo, a través del colículo superior, de un modo análogo al que aplica la corteza visual en los mamíferos.
Este núcleo cumple dos funciones principales: en primer lugar, transfiere la información visual desde el tracto óptico hacia la corteza visual a través de la radiación óptica (también llamada tracto geniculocalcarino). Esta función de relevo es tan precisa que existe una transmisión punto por punto exacta con un importante grado de fidelidad espacial durante todo el trayecto desde la retina hasta la corteza visual. Después de pasar el quiasma, la mitad de las fibras de cada tracto óptico deriva de un ojo y la otra mitad del otro, representando puntos correspondientes de las dos retinas. Sin embargo, los impulsos de ambos ojos se mantienen separados en el núcleo geniculado dorsal lateral. Esta estructura está compuesta por seis capas nucleares. Las capas II, III y V (de ventral a dorsal) reciben señales desde la mitad lateral de la retina del mismo lado, mientras que las capas I, IV y VI las reciben de la mitad medial de la retina del ojo contralateral. Las zonas retinianas respectivas de ambos ojos conectan con neuronas que se encuentran superpuestas en las capas correspondientes y durante todo el trayecto conserva una transmisión paralela semejante hasta la corteza visual. La segunda función más importante del núcleo geniculado dorsal lateral consiste en «filtrar» la transmisión de los impulsos hacia la corteza visual: es decir, controlar qué parte se deja pasar en su camino. El núcleo recibe señales para su acción reguladora de compuerta desde dos fuentes principales: 1) fibras corticófugas que vuelven en sentido retrógrado desde la corteza visual primaria hasta el núcleo geniculado lateral, y 2) las zonas reticulares del mesencéfalo. Estas dos fuentes poseen un carácter inhibidor y, cuando se estimulan, tienen la capacidad de interrumpir la transmisión a través de determinadas porciones del núcleo geniculado dorsal lateral. Estos dos circuitos de filtrado sirven para realzar la información visual que se deja pasar. Finalmente, el núcleo geniculado lateral dorsal está dividido según otro criterio. 1. Las capas I y II se llaman capas magnocelulares porque contienen neuronas grandes. Estas neuronas reciben sus conexiones desde las grandes células ganglionares de la retina de tipo Y casi en su integridad. Este sistema magnocelular proporciona una vía de conducción rápida hacia la corteza visual. Sin embargo, resulta ciego para el color, y no transmite más que información en blanco y negro. Asimismo, su conducción punto por punto es mala porque no hay muchas células ganglionares Y, y sus dendritas ocupan una amplia extensión en la retina. 2. Las capas III a VI se denominan capas parvocelulares porque poseen una gran cantidad de neuronas de tamaño pequeño a mediano. Estas neuronas reciben sus conexiones casi exclusivamente de las células ganglionares retinianas de tipo X que transportan el color y llevan una información espacial precisa punto por punto, pero cuya velocidad de conducción es solo moderada, en vez de alta. Organización y función de la corteza visual Igual que las representaciones corticales de los demás sistemas sensitivos, esta zona está dividida en una corteza visual primaria y áreas visuales secundarias.
Columnas neuronales verticales en la corteza visual La corteza visual posee una organización estructural formada por varios millones de columnas verticales de células neuronales, con un diámetro de 30 a 50 μm cada una. Esta misma disposición en columnas verticales se observa por toda la corteza cerebral dedicada a las demás modalidades sensitivas (y también por las regiones corticales motoras y de análisis). Cada columna constituye una unidad funcional. A grandes rasgos puede calcularse que una columna vertical visual tal vez contenga 1.000 neuronas o más. Una vez que las señales ópticas llegan a la capa IV, sufren una nueva transformación al propagarse hacia el exterior y hacia el interior a lo largo de cada columna vertical. Se cree que este procesamiento descifra componentes independientes de la información visual en las estaciones sucesivas a lo largo de la vía. Los impulsos que ascienden hacia las capas I, II y III acaban enviando su contenido en sentido lateral con un alcance reducido dentro de la corteza. Por el contrario, los que descienden hacia las capas V y VI excitan neuronas que transmiten su actividad a unas distancias mucho mayores «Manchas de color» en la corteza visual Intercaladas entre las columnas visuales primarias y también entre las columnas de algunas áreas visuales secundarias hay unas regiones especiales de tipo columna llamadas manchas de color; estas zonas reciben señales laterales desde las columnas visuales adyacentes y se activan de forma específica por los estímulos de color. Por tanto, se supone que constituyen las zonas primarias para descifrar el color. Interacción de las señales visuales procedentes de ambos ojos Recuerde que las señales visuales procedentes de los dos ojos se transmiten a través de capas neuronales independientes en el núcleo geniculado lateral. Estos impulsos permanecen separados entre sí cuando llegan a la capa IV de la corteza visual primaria. En realidad, la capa IV está entrelazada con las bandas de las columnas neuronales, cuya anchura mide unos 0,5 mm; la actividad procedente de un ojo penetra en las columnas de una banda sí y otra no, según una disposición alterna con las señales del segundo ojo. Este área cortical descifra si las zonas respectivas de las imágenes visuales originadas en cada ojo están «en concordancia» entre sí (es decir, si los puntos correspondientes de las dos retinas coinciden entre ellos). A su vez, la información descifrada se emplea para corregir la mirada direccional de ambos ojos con el fin de que se fusionen entre sí (es decir, se pongan en «concordancia»). Los datos observados sobre el grado de coincidencia entre las imágenes de ambos ojos también permiten que una persona detecte la distancia a un objeto por el mecanismo de la estereopsia.
Dos vías importantes para el análisis de la información visual: 1) la vía rápida de la «posición» y el «movimiento», y 2) la vía de la exactitud del color.
Análisis de la posición tridimensional, la forma global y el movimiento de los objetos – examina la posición tridimensional que ocupan los objetos visuales en el espacio que rodea al organismo. Esta vía también explora la forma física global de la escena visual, así como el movimiento que se produce en su seno. Dicho de otro modo, dice dónde está cada objeto en cada instante y si está en movimiento o no. Después de salir de la corteza visual primaria, los impulsos viajan en general hacia el área temporal media posterior y ascienden hacia la extensa corteza occipitoparietal. En el borde anterior de la corteza parietal se superponen con los que proceden de las áreas de asociación somáticas posteriores dedicadas a analizar los aspectos tridimensionales de las señales somatosensitivas. La información contenida en esta vía encargada de la posición-forma-movimiento básicamente viene de las fibras Y grandes del nervio óptico originadas en las células ganglionares Y de la retina, que transmiten impulsos rápidos pero únicamente de carácter descriptivo en blanco y negro sin color. 2. Análisis de los detalles visuales y del color. Es la vía principal encargada de analizar los detalles visuales. Otras zonas aparte de esta misma vía también se dedican a analizar específicamente el color. Por tanto, esta vía se ocupa de proezas visuales como la identificación de las letras, la lectura, la determinación de la textura de los objetos, de sus colores detallados, y de descifrar lo que es y lo que significa un objeto a partir de toda esta información.
La corteza visual también detecta la orientación de las líneas y los bordes: las células «simples» La corteza visual no solo detecta la existencia de líneas y bordes en las diferentes zonas de la imagen retiniana, sino también la dirección en la que se están orientadas cada una de ellas, es decir, si son verticales u horizontales o si guardan un cierto grado de inclinación. Se cree que esta capacidad obedece a la excitación de neuronas de segundo orden por parte de sistemas lineales de células que se inhiben mutuamente, cuando esta inhibición se produce siguiendo toda la línea celular que corresponde a un límite de contraste. Por tanto, con cada orientación de una línea de este tipo, se estimulan células neuronales específicas. Una línea dispuesta según una dirección diferente excita un grupo de células distinto. Estas neuronas se denominan células simples. Sobre todo están situadas en la capa IV de la corteza visual primaria. Detección de la orientación lineal cuando una línea se desplaza en sentido lateral o vertical a lo largo del campo visual: las células «complejas» A medida que la señal visual se aleja de la capa IV, algunas neuronas responden a líneas orientadas en la misma dirección pero sin ser específicas de una posición. Esto es, incluso si una línea recorre por el campo una distancia moderada en sentido lateral o vertical, estas mismas neuronas poco abundantes aún seguirán estimuladas si la línea conserva la misma dirección. Dichas neuronas se llaman células complejas. Detección de líneas con longitudes, ángulos u otras formas específicos Algunas neuronas de las capas más externas de las columnas visuales primarias, lo mismo que las pertenecientes a ciertas áreas visuales secundarias, no se estimulan más que por líneas o bordes con una longitud o una forma en ángulo específicas, o por imágenes que poseen otras características. Es decir, estas neuronas detectan órdenes aún más elevadas de información procedentes de la escena visual. Por tanto, a medida que uno asciende dentro de la vía analítica de la corteza visual, se descifran cada vez más características de una escena visual. Detección del color El color se detecta básicamente por el mismo procedimiento que las líneas: mediante su contraste. Por ejemplo, una zona roja suele quedar contrastada frente a otra verde, una azul contra una roja, o una verde contra una amarilla. Todos estos colores también pueden compararse frente a una zona blanca en el seno de la escena visual. De hecho, se piensa que este contraste con el blanco es el responsable principal del fenómeno llamado «constancia del color»; a saber, cuando el color de un haz de iluminación cambia, también lo hace el tono del «blanco» con esta luz, y el cálculo oportuno por parte del cerebro permite interpretar el rojo como rojo aun cuando el haz de iluminación haya modificado el color que llega a los ojos. El mecanismo para analizar el contraste de color depende del hecho de que los tonos de contraste, llamados «colores contrarios», excitan células neuronales específicas. Se supone que las células simples detectan los primeros detalles para el contraste de color, mientras que los más complicados están a cargo de las células complejas e hipercomplejas.
Anomalías del campo visual A veces, los puntos ciegos se encuentran en porciones del campo visual diferentes al área del disco óptico. Tales puntos, llamados escotomas, a menudo están causados por el daño del nervio óptico como consecuencia de un glaucoma (presión excesiva del líquido contenido en el interior del globo ocular), las reacciones alérgicas a nivel de la retina o procesos tóxicos, como el saturnismo o el consumo excesivo de tabaco. Otro trastorno que puede diagnosticarse con la campimetría es la retinitis pigmentaria. En esta enfermedad, ciertas porciones de la retina degeneran, y en las zonas degeneradas se deposita un exceso del pigmento melanina. La retinitis pigmentaria primero suele ocasionar una ceguera en el campo visual periférico y a continuación invadir gradualmente las áreas centrales. Efecto de las lesiones de la vía óptica sobre los campos visuales La destrucción de todo el nervio óptico origina una ceguera en el ojo afectado. Si el trastorno afecta al quiasma óptico impide el cruce de los impulsos que proceden de la mitad nasal de cada retina y van dirigidos hacia el tracto óptico del lado opuesto. Por tanto, esta mitad queda ciega a ambos lados, lo que significa que la persona está ciega en el campo temporal de cada ojo porque la imagen del campo visual se encuentra invertida en la retina debido al sistema óptico ocular; esta situación se denomina hemianopsia bitemporal. Tales lesiones normalmente son el resultado de un tumor hipofisario que presiona sobre la parte inferior del quiasma desde la silla turca hacia arriba. La interrupción de un tracto óptico deja sin inervación la mitad de cada retina correspondiente al mismo lado de la lesión; como consecuencia, ningún ojo es capaz de ver los objetos situados en el lado opuesto de la cabeza. Dicha situación se conoce como hemianopsia homónima. Movimientos oculares y su control Para sacar todo el partido a las capacidades visuales de los ojos, el sistema de control cerebral encargado de dirigirlos hacia el objeto que pretende observarse tiene casi tanta importancia como la interpretación de las señales visuales nacidas en ellos.
Movimientos oculares de fijación Tal vez los movimientos más importantes de los ojos son los que provocan su «fijación» en una porción concreta del campo visual. Este fenómeno está controlado por dos mecanismos neuronales. El primero de estos mecanismos permite a una persona mover los ojos voluntariamente para encontrar el objeto sobre el que desea fijar la visión, que se llama mecanismo voluntario de fijación. El segundo es un proceso involuntario, llamado mecanismo involuntario de fijación, que mantiene los ojos fijos con firmeza sobre el objeto una vez que ha sido descubierto. Mecanismo de bloqueo involuntario de la fijación: función de los colículos superiores El tipo de bloqueo involuntario de la fijación comentado en el apartado anterior resulta de un mecanismo de retroalimentación negativo encargado de evitar que el objeto de atención se salga de la porción retiniana correspondiente a la fóvea. Los ojos normalmente poseen tres tipos de movimientos constantes, pero casi imperceptibles: 1) un temblor continuo a una frecuencia de 30 a 80 ciclos/s ocasionado por las contracciones sucesivas de las unidades motoras en los músculos oculares; 2) una lenta traslación de los globos oculares en una dirección u otra, y 3) movimientos de sacudida súbitos que están controlados por el mecanismo involuntario de fijación.
Movimientos sacádicos de los ojos: un mecanismo formado por puntos sucesivos de fijación Si una escena visual sufre un desplazamiento continuo delante de los ojos, como sucede cuando una persona va en coche, estos irán fijándose en los elementos más destacados del campo visual uno tras otro, saltando desde cualquiera de ellos al siguiente a una velocidad de dos a tres saltos por segundo. Estos saltos se llaman sacadas y los movimientos se denominan movimientos optocinéticos. Las sacadas ocurren a tal velocidad que el movimiento de los ojos no ocupa ni el 10% del tiempo total y el 90% queda dedicado a los lugares de fijación. Asimismo, el cerebro suprime la visión de la imagen durante las sacadas, de modo que la persona no tiene conciencia de los movimientos realizados de un punto a otro. Movimientos sacádicos durante la lectura Durante el proceso de lectura, una persona suele realizar varios movimientos sacádicos oculares en cada línea. En este caso, la escena visual no se está moviendo delante de los ojos, pero estos están entrenados para desplazarse por medio de varias sacadas sucesivas a través de la escena visual con el fin de extraer la información importante. Otras sacadas semejantes ocurren cuando una persona observa un cuadro excepto que en esta ocasión las sacudidas se suceden una tras otra hacia arriba, hacia abajo, hacia los lados y siguiendo trayectorias en ángulo desde un aspecto sobresaliente a otro, y así sucesivamente. Fijación de objetos en movimiento: «movimiento de seguimiento» Los ojos también pueden permanecer fijos sobre un objeto que se esté desplazando, lo que se denomina movimiento de seguimiento. Un mecanismo cortical muy avanzado detecta automáticamente la trayectoria seguida por el objeto en movimiento y a continuación concibe con rapidez un recorrido similar para el desplazamiento de los ojos. Por ejemplo, si un objeto sube y baja en forma de onda a una frecuencia de varias oscilaciones por segundo, al principio los ojos pueden ser incapaces de fijarlo en la mirada. Sin embargo, pasado 1 s más o menos, comienzan a saltar en virtud de las sacadas aproximadamente con el mismo patrón de movimiento ondulatorio que el del objeto. A continuación, después de unos cuantos segundos más, los ojos adoptan unos movimientos cada vez más suaves y finalmente siguen el desplazamiento de la onda casi con absoluta exactitud. Esto representa un gran nivel de capacidad automática inconsciente de cálculo por parte del sistema de seguimiento encargado de controlar los movimientos oculares. Los colículos superiores son los principales responsables del giro de los ojos y de la cabeza para dirigirlos hacia una perturbación visual Incluso después de haber desaparecido la corteza visual, cualquier perturbación repentina en una zona lateral del campo visual suele suscitar el giro inmediato de los ojos en esa dirección. Este giro no sucede si también se han destruido los colículos superiores. Para cumplir esta función existe una representación topográfica de los diversos puntos de la retina en estas últimas estructuras de un modo parecido a lo que sucede en la corteza visual primaria, aunque con menor exactitud. Aun así, la orientación principal de un destello luminoso en el campo periférico de la retina viene cartografiada por los colículos, que transmiten señales secundarias hacia los núcleos oculomotores con el fin de girar los ojos. Para colaborar con este movimiento ocular direccional, los colículos superiores también poseen mapas topográficos de las sensaciones somáticas originadas en el cuerpo y de las acústicas procedentes del oído. Las fibras del nervio óptico que van desde los ojos hasta los colículos y son las responsables de estos movimientos rápidos de giro son ramas de las fibras M de conducción rápida, uno de cuyos extremos se dirige hacia la corteza visual y el otro hacia los colículos superiores. Además de hacer que los ojos se vuelvan hacia una perturbación visual, las señales se transfieren desde los colículos superiores hacia otros niveles del tronco del encéfalo a través del fascículo longitudinal medial para provocar el giro de toda la cabeza e incluso de todo el cuerpo en esa misma dirección. Otros tipos de acontecimientos no visuales, como los sonidos potentes o incluso un golpe recibido en el costado del cuerpo, provocan un giro similar de los ojos, la cabeza y el tronco, pero solo si los colículos superiores están íntegros. Por tanto, estas estructuras cumplen un cometido global en la orientación de los ojos, la cabeza y el cuerpo con respecto a las perturbaciones externas, sean de carácter visual, auditivo o somático. «Fusión» de las imágenes visuales de ambos ojos Para que las percepciones visuales resulten más elocuentes, normalmente se fusionan entre sí las imágenes visuales de ambos ojos según los «puntos correspondientes» de las dos retinas. La corteza visual ocupa un lugar importante en el proceso de fusión. Ya se indicó antes en este mismo capítulo que los puntos correspondientes de ambas retinas transmiten señales visuales hacia capas neuronales diferentes en el cuerpo geniculado lateral y a su vez estos impulsos se transfieren hasta las neuronas situadas en una posición paralela en la corteza visual. Estas neuronas corticales interaccionan entre sí, dando lugar a la excitación por interferencia de neuronas específicas cuando las dos imágenes visuales no quedan «en concordancia», es decir, cuando su «fusión» no se realiza con precisión. Se supone que esta excitación suministra la señal que se transmite al aparato oculomotor para provocar la convergencia, la divergencia o la rotación de los ojos a fin de que pueda restablecerse la fusión. Una vez que coinciden los puntos correspondientes de las dos retinas, desaparece la excitación de las neuronas específicas «de interferencia» en la corteza visual.
Estrabismo: falta de fusión de los ojos El estrabismo, también denominado bizquera o desviación de los ojos, quiere decir la falta de fusión entre los ojos en una coordenada visual o más: la horizontal, la vertical o la de rotación.
Supresión de la imagen visual procedente de un ojo reprimido En unos pocos pacientes con estrabismo, el ojo que se fija sobre el objeto de atención sufre un proceso de alternancia. Otros no emplean más que un ojo todo el tiempo, y el contrario queda reprimido y nunca se utiliza para la visión con detalle. La agudeza visual del ojo reprimido solo se desarrolla ligeramente, y a veces se queda en 20/400 o menos. Si el ojo dominante más tarde sufre una ceguera, la visión del ojo reprimido únicamente puede desplegarse hasta cierto punto en los adultos, pero mucho más en los niños pequeños. Esto pone de manifiesto que la agudeza visual depende en gran medida de la correcta formación de las conexiones sinápticas oculares en el sistema nervioso central. En realidad, incluso a escala anatómica, el número de conexiones neuronales disminuye en las áreas de la corteza visual que normalmente recibirían señales desde el ojo reprimido. NERVIOS AUTONOMOS DEL OJO. Las fibras preganglionares parasimpáticas nacen en el núcleo de Edinger-Westphal (la porción nuclear visceral del tercer par craneal) y a continuación viajan en el tercer par hasta el ganglio ciliar, que se halla justo detrás del ojo. En este punto, los axones preganglionares hacen sinapsis con las neuronas parasimpáticas posganglionares, que a su vez envían sus fibras hacia el globo ocular a través de los nervios ciliares. Estos nervios excitan: 1) el músculo ciliar que controla el enfoque del cristalino, y 2) el esfínter del iris que con La inervación simpática del ojo se origina en las células del asta intermediolateral a nivel del primer segmento torácico de la médula espinal. Desde allí, las fibras simpáticas penetran en la cadena simpática y ascienden hacia el ganglio cervical superior, donde realizan su sinapsis con las neuronas posganglionares. Las fibras simpáticas posganglionares siguen a continuación desde aquí a lo largo de la superficie de la arteria carótida y de otras arterias cada vez más pequeñas hasta que llegan al ojo. En esta estructura, inervan las fibras radiales del iris (que abren la pupila) así como varios músculos extraoculares, que se comentan más adelante en relación con el síndrome de Horner
La acomodación del cristalino está regulada por un mecanismo de retroalimentación negativo que corrige automáticamente su poder dióptrico para lograr el mayor grado de agudeza visual. Cuando los ojos se han enfocado en algún objeto lejano y a continuación deben cambiar bruscamente para captar otro objeto próximo, el cristalino suele acomodarse para conseguir la mejor agudeza posible de la visión en menos de 1 s. Aunque no está claro cuál es el mecanismo de control exacto que procura este enfoque rápido y preciso del ojo, se conocen las siguientes características. En primer lugar, cuando los ojos modifican repentinamente la distancia de su punto de fijación, el cristalino cambia su potencia de la forma pertinente para alcanzar un nuevo estado de enfoque en cuestión de una fracción de segundo. En segundo lugar, diversos tipos de datos sirven para transformar la potencia del cristalino en el sentido apropiado: 1. La aberración cromática parece un elemento importante. Es decir, los rayos de luz rojos se enfocan un poco más atrás que los azules debido a que el cristalino provoca una desviación de estos últimos superior a la de aquellos. El ojo parece ser capaz de detectar cuál de estos dos tipos de rayos está mejor enfocado y este dato transmite información hacia el mecanismo de acomodación con el fin de aumentar o reducir la potencia del cristalino. 2. Cuando los ojos se fijan sobre un objeto cercano, deben converger. Los mecanismos nerviosos de la convergencia generan una señal simultánea para aumentar la potencia del cristalino. 3. Dado que la fóvea se halla situada en una depresión hueca que queda un poco más honda que el resto de la retina, la claridad de enfoque en su profundidad es diferente de la claridad de enfoque en los bordes. Esta diferencia también aporta una pista sobre el sentido en el que resulta necesario modificar la potencia del cristalino. 4. El grado de acomodación del cristalino oscila un poco todo el tiempo a una frecuencia que llega hasta dos cambios por segundo. La imagen visual gana claridad cuando la oscilación de su potencia sigue el sentido adecuado y la pierde cuando lleva el sentido erróneo. Esto podría aportar un indicio rápido para decidir hacia dónde ha de corregirse esta potencia si se quiere conseguir el enfoque pertinente. Las áreas corticales cerebrales dedicadas a controlar la acomodación siguen fielmente las que se encargan de los movimientos oculares de fijación. El análisis de las señales visuales en las áreas corticales 18 y 19 de Brodmann y la transmisión de las señales motoras hacia el músculo ciliar tienen lugar a través del área pretectal en el tronco del encéfalo, para seguir después por el núcleo de Edinger-Westphal y finalmente alcanzar los ojos por medio de las fibras nerviosas parasimpáticas.
Reflejos o reacciones pupilares en las enfermedades del sistema nervioso central Unas cuantas enfermedades del sistema nervioso central dañan la transmisión nerviosa de señales visuales desde la retina hasta el núcleo de Edinger-Westphal, lo que a veces acaba con los reflejos pupilares. Este bloqueo puede ocurrir como consecuencia de una sífilis del sistema nervioso central, el alcoholismo, una encefalitis, etc. Normalmente sucede en la región pretectal del tronco del encéfalo, aunque puede obedecer a la destrucción de ciertas fibras pequeñas en los nervios ópticos. Las fibras nerviosas finales de la vía que atraviesa el área pretectal en su camino hasta el núcleo de Edinger-Westphal poseen en su mayoría un carácter inhibidor. Cuando su efecto desaparece, el núcleo queda activo de forma prolongada, lo que se traduce en que las pupilas permanezcan básicamente contraídas, además de que no respondan a la luz. Con todo, las pupilas pueden contraerse un poco más si el núcleo de Edinger-Westphal recibe un estímulo por cualquier otra vía. Por ejemplo, cuando los ojos se fijan en un objeto cercano, las señales que provocan la acomodación del cristalino y las que causan la convergencia de ambos ojos generan al mismo tiempo un pequeño grado de contracción pupilar. Este fenómeno se denomina reacción pupilar a la acomodación. Una pupila que no responda a la luz, pero sí a la acomodación y cuyo tamaño es muy pequeño (la pupila de Argyll Robertson) es un signo diagnóstico importante de una enfermedad en el sistema nervioso central, por ejemplo de una sífilis. Síndrome de Horner Los nervios simpáticos del ojo a veces quedan interrumpidos. Esta circunstancia suele suceder en la cadena simpática cervical, lo que provoca el cuadro clínico llamado síndrome de Horner. Este síndrome ocasiona los siguientes efectos. Primero, debido a la interrupción de las fibras nerviosas simpáticas dirigidas al músculo dilatador de la pupila, esta permanece contraída de forma continua con un diámetro más pequeño que la pupila del lado opuesto. Segundo, el párpado superior se cae debido a que normalmente se mantiene en posición abierta durante las horas de vigilia en parte por la contracción de las fibras musculares lisas contenidas en su interior e inervadas por el sistema simpático. Por tanto, la destrucción de estos nervios imposibilita su apertura hasta una altura normal. Tercero, los vasos sanguíneos del lado correspondiente de la cara y de la cabeza quedan dilatados de un modo persistente. Cuarto, no puede producirse la sudoración (que requiere la acción de las señales nerviosas simpáticas) en el mismo lado de la cara y de la cabeza afectado por el síndrome de Horner.
La sensibilidad somática es el mecanismo nervioso que recopila la información sensitiva de todo el cuerpo. Estos sentidos se contraponen a las sensibilidades especiales, que aluden específicamente a la vista, el oído, el olfato, el gusto y el equilibrio.
Clasificación de las sensibilidades somáticas. Las sensibilidades somáticas pueden clasificarse en tres tipos fisiológicos: 1) las sensibilidades somáticas mecanorreceptoras, formadas por las sensaciones táctiles y posicionales cuyo estímulo depende del desplazamiento mecánico de algún tejido del organismo 2) las sensibilidades termorreceptoras, que detectan el calor y el frío 3) la sensibilidad al dolor, que se activa con factores que dañan los tejidos. OTRAS CLASIFICACIONES DE LAS SENSIBILIDADES SOMATICAS. La sensibilidad exterorreceptora: es la que procede de la superficie del cuerpo. La sensibilidad propioceptiva: es la que tiene que ver con el estado físico del cuerpo, como las sensaciones posicionales, las tendinosas y musculares, las de presión originadas en la planta de los pies e incluso la sensación de equilibrio (que a menudo se considera una sensibilidad «especial» en vez de una sensibilidad somática). La sensibilidad visceral: es la que deriva de las vísceras del cuerpo; al emplear este término, uno suele referirse en concreto a las sensaciones de los órganos internos. La sensibilidad profunda: es la que viene de los tejidos profundos, como las fascias, los músculos y los huesos, esta sensibilidad comprende básicamente la presión «profunda», el dolor y la vibración.
Detección y transmisión de las sensaciones táctiles. Aunque el tacto, la presión y la vibración suelen clasificarse como sensaciones independientes, todas se detectan mediante los mismos tipos de receptores. Existen tres diferencias principales entre ellas: 1) la sensación de tacto en general deriva de la estimulación de los receptores táctiles situados en la piel o en los tejidos inmediatamente por debajo de ella 2) la sensación de presión suele obedecer a la deformación de los tejidos profundos, 3) la sensación de vibración resulta de la repetición de señales sensitivas con rapidez, pero recurre a algunos tipos de receptores que también emplean las de tacto y de presión. RECEPTORES TÁCTILES. En primer lugar, algunas terminaciones nerviosas libres, que están distribuidas por todas partes en la piel y en otros muchos tejidos, son capaces de detectar el tacto y la presión. Por ejemplo, incluso un contacto suave con la córnea, que no contiene ningún otro tipo de terminaciones nerviosas aparte de las libres, puede llegar a despertar sensaciones de tacto y de presión. En segundo lugar un segundo receptor dotado es el corpúsculo de Meissner. que es una terminación nerviosa encapsulada alargada perteneciente a una gran fibra nerviosa sensitiva mielínica (de tipo Aβ). Dentro de la cápsula hay muchos filamentos nerviosos terminales ramificados. Estos corpúsculos están presentes en las partes de la piel desprovistas de pelo o lampiñas y son especialmente abundantes en las yemas de los dedos, en los labios y en otras zonas cutáneas que presenten una capacidad muy desarrollada para discernir la localización espacial de las sensaciones táctiles. Los corpúsculos de Meissner se adaptan en cuestión de una fracción de segundo después de ser estimulados, lo que significa que resultan especialmente sensibles al movimiento de los objetos sobre la superficie de la piel, y también a la vibración de baja frecuencia. En tercer lugar, las yemas de los dedos y otras zonas que contienen una gran cantidad de corpúsculos de Meissner también suelen albergar un número elevado de receptores táctiles de terminación bulbar, uno de cuyos ejemplos son los discos de Merkel. Las partes de la piel cubiertas de pelo presentan asimismo una cuantía moderada de receptores de terminación bulbar, aunque prácticamente carecen de corpúsculos de Meissner. Estas estructuras difieren de los corpúsculos de Meissner porque al principio transmiten una señal intensa pero parcialmente adaptable, y después una señal más débil y continua que se adapta solo con lentitud. Por tanto, son responsables de suministrar las señales estables que permiten determinar un contacto continuo de los objetos contra la piel.
Transmisión de señales táctiles en las fibras nerviosas periféricas. Casi todos los receptores sensitivos especializados, como los corpúsculos de Meissner, los receptores en cúpula de Iggo, los receptores pilosos, los corpúsculos de Pacini y las terminaciones de Ruffini, envían sus señales por fibras nerviosas de tipo Aβ que poseen una velocidad de transmisión entre 30 y 70 m/s. Por el contrario, los receptores táctiles de las terminaciones nerviosas libres mandan sus señales sobre todo a través de pequeñas fibras mielínicas de tipo Aδ que no conducen más que a una velocidad de 5 a 30 m/s. Algunas terminaciones nerviosas libres para el tacto recurren a fibras amielínicas de tipo C cuyas velocidades oscilan desde mucho menos de 1 m hasta 2 m/s; estas terminaciones nerviosas envían las señales hacia la médula espinal y la parte inferior del tronco del encéfalo, probablemente sobre todo al servicio de la sensación de cosquilleo. Detección de la vibración Todos los receptores táctiles participan en la detección de las vibraciones, aunque los distintos receptores detectan frecuencias también distintas. Los corpúsculos de Pacini pueden identificar vibraciones con señales desde 30 hasta 800 ciclos/s debido a que responden con una rapidez extrema a las deformaciones minúsculas y veloces de los tejidos. También envían sus señales a las fibras nerviosas de tipo Aβ, que son capaces de transmitir hasta 1.000 impulsos por segundo. Por el contrario, las vibraciones de baja frecuencia, desde 2 hasta 80 ciclos/s. estimulan otros receptores táctiles, sobre todo los corpúsculos de Meissner, cuya adaptación es más lenta que en los corpúsculos de Pacini. Detección del cosquilleo y el picor por terminaciones nerviosas libres mecanorreceptoras. Los estudios neurofisiológicos han demostrado la existencia de terminaciones nerviosas libres mecanorreceptoras de adaptación rápida y muy sensibles que tan solo suscitan sensaciones de cosquilleo y de picor. Asimismo, estas terminaciones se encuentran casi exclusivamente en las capas superficiales de la piel, que también es el único tejido desde el que suelen poder despertarse estos fenómenos. Dichas sensaciones se transmiten por fibras amielínicas muy pequeñas de tipo C semejantes a las que se encargan del dolor de tipo lento y continuo. Vías sensitivas para la transmisión de señales somáticas en el sistema nervioso central. Casi toda la información sensitiva procedente de los segmentos somáticos corporales penetra en la médula espinal a través de las raíces dorsales de los nervios raquídeos. Sin embargo, desde su punto de entrada estas señales son transmitidas por la médula y más tarde por el encéfalo a través de una de las dos vías sensitivas alternativas siguientes: 1) el sistema de la columna dorsal-lemnisco medial, o 2) el sistema anterolateral. Estos dos caminos vuelven a reunirse parcialmente a nivel del tálamo. El sistema de la columna dorsal-lemnisco medial, como su nombre da a entender, transporta señales en sentido ascendente básicamente por las columnas dorsales de la médula hacia el bulbo raquídeo en el encéfalo. A continuación, después de hacer sinapsis y cruzar al lado opuesto a este nivel, siguen subiendo a través del tronco del encéfalo hasta el tálamo dentro del lemnisco medial. Por el contrario, las señales del sistema anterolateral, nada más entrar en la médula espinal procedentes de las raíces dorsales de los nervios raquídeos, hacen sinapsis en las astas dorsales de la sustancia gris medular, después cruzan al lado opuesto y ascienden a través de sus columnas blancas anterior y lateral. Su terminación se produce a todos los niveles de la parte inferior del tronco del encéfalo y en el tálamo. El sistema de la columna dorsal-lemnisco medial está compuesto por fibras nerviosas mielínicas grandes que transmiten señales hacia el cerebro a una velocidad de 30 a 110 m/s, mientras que el sistema anterolateral está integrado por fibras mielínicas más pequeñas cuya velocidad de transmisión oscila desde unos pocos metros por segundo hasta 40 m/s.
Transmisión por el sistema de la columna dorsallemnisco medial. Anatomía del sistema de la columna dorsal-lemnisco medial. Al penetrar en la médula espinal a través de las raíces dorsales de los nervios raquídeos, las grandes fibras mielínicas procedentes de los mecanorreceptores especializados se dividen casi de inmediato para dar lugar a una rama medial y una rama lateral.
Orientación espacial de las fibras nerviosas en el sistema de la columna dorsal-lemnisco medial. Uno de los rasgos diferenciadores del sistema de la columna dorsal-lemnisco medial es la diversa orientación espacial que adquieren las fibras nerviosas procedentes de cada parte del cuerpo, y que se mantiene todo el tiempo. Por ejemplo, en las columnas dorsales de la médula espinal, las fibras de las porciones inferiores del organismo quedan situadas hacia el centro de la médula, mientras que las que entran en ella a niveles segmentarios paulatinamente superiores forman capas sucesivas más laterales. En el tálamo, esta orientación espacial distinta aún se conserva, estando representado el extremo caudal del cuerpo en las porciones más laterales del complejo ventrobasal y la cabeza y la cara en sus zonas mediales. Debido al cruce que experimentan los lemniscos en el bulbo raquídeo, el lado izquierdo del cuerpo se representa a la derecha en el tálamo y el derecho a la izquierda. CORTEZA SOMATOSENSITIVA.
Áreas estructuralmente distintas de la corteza cerebral humana, llamadas áreas de Brodmann. Obsérvense específicamente las áreas 1, 2 y 3, que constituyen el área somatosensitiva primaria I, y las áreas 5 y 7A, que constituyen el área de asociación somatosensitiva, la gran cisura central (también llamada surco central) que se extiende en sentido horizontal cruzando el cerebro. En general, las señales sensitivas pertenecientes a cualquier modalidad de sensación terminan en la corteza cerebral inmediatamente por detrás de la cisura central. También a grandes rasgos, la mitad anterior del lóbulo parietal se ocupa casi por completo de la recepción e interpretación de las señales somatosensitivas, pero la mitad posterior aporta unos niveles aún más altos de interpretación. Las señales visuales acaban en el lóbulo occipital, y las señales auditivas terminan en el lóbulo temporal. Por el contrario, la porción de la corteza cerebral que queda delante de la cisura central y constituye la mitad posterior del lóbulo frontal se llama corteza motora y está dedicada casi en su integridad a controlar las contracciones musculares y los movimientos del cuerpo. Un ingrediente principal de este control motor llega en respuesta a las señales somatosensitivas recibidas desde las porciones corticales sensitivas, que mantienen informada a cada instante a la corteza motora sobre las posiciones y los movimientos de las diferentes partes del cuerpo. Áreas somatosensitivas I y II El área somatosensitiva I presenta un grado acusado de localización de las diferentes porciones corporales, en cambio, el grado de localización es escaso en el área somatosensitiva II, aunque, a grandes rasgos, la cara está representada en su zona anterior, los brazos en la central y las piernas en la posterior. Mucho menos se sabe acerca de la función del área somatosensitiva II. Sí se ha observado que las señales llegan a ella desde el tronco del encéfalo, transmitidas en sentido ascendente a partir de las dos mitades del cuerpo. Además, un origen secundario de muchas de ellas está en el área somatosensitiva I, lo mismo que en otras áreas sensitivas del cerebro, incluso las visuales y auditivas. Son necesarias las proyecciones desde el área somatosensitiva I para que el área somatosensitiva II funcione. Sin embargo, la extirpación de una parte carece de efectos aparentes sobre la respuesta de las neuronas pertenecientes al área somatosensitiva I. Así pues, mucho de lo que sabemos sobre la sensibilidad somática parece explicarse por las funciones del área somatosensitiva I. El área somatosensitiva I se halla inmediatamente detrás de la cisura central, situada en la circunvolución poscentral de la corteza cerebral humana (corresponde a las áreas de Brodmann 3, 1 y 2).
Capas de la corteza somatosensitiva y su función La corteza cerebral contiene seis capas de neuronas, comenzando por la capa I próxima a la superficie cerebral y siguiendo cada vez por zonas más profundas hasta la capa VI. 1.La señal sensitiva entrante excita en primer lugar la capa neuronal IV; a continuación, se propaga hacia la superficie de la corteza y también hacia otras capas más profundas. 2. Las capas I y II reciben señales de entrada difusas inespecíficas procedentes de los centros inferiores del encéfalo, que facilitan regiones corticales específicas. 3.Las neuronas de las capas II y III envían axones hacia las porciones emparentadas entre sí de la corteza cerebral en el lado opuesto del cerebro a través del cuerpo calloso. 4. Las neuronas de las capas V y VI mandan axones hacia las partes más profundas del sistema nervioso. Las de la capa V en general son mayores y proyectan hacia zonas más alejadas, como los ganglios basales, el tronco del encéfalo y la médula espinal, donde controlan la transmisión de la señal. Desde la capa VI, un número especialmente grande de axones se extiende hacia el tálamo, suministrando señales corticales, que interaccionan con las señales sensitivas de entrada que llegan al tálamo, y sirven para regular sus niveles excitadores.
La corteza sensitiva está organizada en columnas verticales de neuronas; cada columna detecta un lugar sensitivo diferente en el cuerpo con una modalidad sensitiva específica Desde el punto de vista funcional, las neuronas de la corteza somatosensitiva están dispuestas formando columnas verticales que se extienden a lo largo de las seis capas corticales, con un diámetro de 0,3 a 0,5 mm y un contenido quizá de 10.000 somas neuronales. Cada una de estas columnas se dedica a una sola modalidad sensitiva específica: algunas columnas responden a los receptores de estiramiento que rodean a las articulaciones, otras a la estimulación de los receptores táctiles pilosos, o a los distintos puntos de presión localizados en la piel, etc. En la capa IV, donde llegan primero las señales sensitivas de entrada a la corteza, las columnas neuronales funcionan casi separadas por completo una de otra. A otros niveles, se producen interacciones que sirven para iniciar el análisis de los significados portados por las señales sensitivas. En los 5 a 10 mm más anteriores de la circunvolución poscentral, situados en el área 3A de Brodmann en la profundidad de la cisura central, una porción especialmente grande de las columnas verticales responde a los receptores de estiramiento articulares, tendinosos y musculares. Muchas de las señales procedentes de estas columnas sensitivas se difunden después en sentido anterior, directamente a la corteza motora localizada justo por delante de la cisura central; así, desempeñan un papel fundamental en el control de las señales motoras de salida que activan secuencias de contracción muscular. A medida que uno se aleja hacia atrás en el área somatosensitiva I, las columnas verticales responden cada vez más a los receptores cutáneos de adaptación lenta, y yendo todavía más hacia atrás, crece el número de columnas sensibles a la presión profunda. En la porción más posterior del área somatosensitiva I, aproximadamente el 6% de las columnas verticales responde solo cuando un estímulo se desplaza a través de la piel en una dirección particular. Por tanto, este es un orden de interpretación de las señales sensitivas aún superior; el proceso se vuelve todavía más complejo cuando las señales se diseminan más lejos en sentido posterior desde el área somatosensitiva I hacia la corteza parietal, una zona llamada área de asociación somatosensitiva, según comentamos más adelante. Funciones del área somatosensitiva I La resección bilateral generalizada del área somatosensitiva I provoca la desaparición de los siguientes tipos de evaluación sensitiva: 1. La persona es incapaz de localizar las diversas sensaciones de forma diferenciada en las distintas partes del cuerpo. Sin embargo, sí puede hacerlo de un modo rudimentario, como en una mano concreta, en un gran nivel del tronco o en una de las piernas. Por tanto, está claro que el tronco del encéfalo, el tálamo o porciones de la corteza que normalmente no se consideran relacionadas con la sensibilidad somática pueden lograr cierto grado de localización. 2. La persona es incapaz de valorar un grado crítico de presión sobre el cuerpo. 3. La persona es incapaz de valorar el peso de los objetos. 4. La persona es incapaz de valorar las formas o la configuración de los objetos. Este trastorno se llama astereognosia. 5. La persona es incapaz de valorar la textura de los materiales porque este tipo de evaluación depende de sensaciones muy críticas originadas por el movimiento de los dedos sobre la superficie que se pretende explorar. Obsérvese que en la lista no se dice nada sobre la desaparición de la sensibilidad al dolor y la temperatura. Ante una ausencia específica solo del área somatosensitiva I, aún se conserva la apreciación de estas modalidades sensitivas en lo que atañe a su cualidad y su intensidad.
Sensibilidades posicionales. Las sensibilidades posicionales también se denominan a menudo sensibilidades propioceptivas y pueden dividirse en dos subtipos: 1) sensibilidad posicional estática, que significa la percepción consciente de la orientación de las diferentes partes del cuerpo unas respecto a otras 2) velocidad de la sensibilidad al movimiento, también llamada cinestesia o propiocepción dinámica. Receptores sensitivos posicionales El conocimiento de la posición, tanto estática como dinámica, depende de la información sobre el grado de angulación de todas las articulaciones en cualquiera de los planos y sus velocidades de cambio. Por tanto, son múltiples los diferentes tipos de receptores que sirven para determinar la angulación articular y que se emplean en conjunto dentro de la sensibilidad posicional. Intervienen tanto receptores táctiles cutáneos como receptores profundos cercanos a las articulaciones. En el caso de los dedos de la mano, donde los receptores cutáneos son muy abundantes, se cree que hasta la mitad de la identificación posicional depende de su detección. A la inversa, en la mayoría de las articulaciones grandes del cuerpo, los receptores profundos cobran mayor trascendencia. Entre los receptores más relevantes que sirven para determinar la angulación articular en el recorrido medio del movimiento figuran los husos musculares.
Algunos aspectos especiales del funcionamiento somatosensitivo Función del tálamo en la sensibilidad somática Cuando se destruye la corteza somatosensitiva de un ser humano, esa persona pierde las sensibilidades táctiles más críticas, pero recupera un ligero grado de sensibilidad táctil grosera. Por tanto, debe suponerse que el tálamo (lo mismo que otros centros inferiores) posee una pequeña capacidad de distinguir las sensaciones táctiles, aun cuando normalmente se dedica sobre todo a transmitir este tipo de información hacia la corteza. En cambio, la desaparición de la corteza somatosensitiva ejerce un efecto escaso sobre la percepción individual de las sensaciones dolorosas y solo un efecto moderado sobre la percepción de la temperatura. Por tanto, la parte baja del tronco del encéfalo, el tálamo y otras regiones basales del encéfalo emparentadas con ellas representan un papel dominante en el discernimiento de estas sensibilidades. Es interesante que dichas modalidades aparezcan muy pronto en el curso del desarrollo filogénico de los animales, mientras que las sensibilidades táctiles críticas y la corteza somatosensitiva sean fenómenos tardíos. Control cortical de la sensibilidad sensitiva: señales «corticófugas» Además de la información somatosensitiva transmitida desde la periferia hacia el cerebro, las señales corticófugas siguen un sentido retrógrado desde la corteza cerebral hacia las estaciones de relevo sensitivo inferiores en el tálamo, el bulbo raquídeo y la médula espinal; se encargan de controlar la intensidad de la sensibilidad que presentan las entradas sensitivas. Las señales corticófugas tienen un carácter inhibidor casi en su integridad, de modo que cuando la intensidad de la entrada sensitiva adquiere demasiado volumen, su intervención reduce la transmisión automáticamente en los núcleos de relevo. Esta acción genera dos efectos: en primer lugar, disminuye la dispersión lateral de las señales sensitivas hacia las neuronas adyacentes y, por tanto, acentúa el grado de nitidez en el patrón de la señal. En segundo lugar, mantiene al sistema sensitivo operando dentro de unos márgenes de sensibilidad que no son tan bajos como para que las señales resulten inútiles ni tan altos como para que el sistema quede anegado por encima de su capacidad para diferenciar los patrones sensitivos. Este principio del control sensitivo corticófugo lo emplean todos los sistemas sensitivos, no solo el somático.
Sensibilidades somáticas: II. Dolor, cefalea y sensibilidad térmica. Muchas de las dolencias del cuerpo generan dolor. Además, la capacidad para diagnosticar las diferentes enfermedades depende en gran medida de los conocimientos que posea un médico sobre las diversas cualidades del dolor. Por estas razones, la primera parte de este capítulo está dedicada básicamente al dolor y a los fundamentos fisiológicos de algunos fenómenos clínicos asociados a él. El dolor aparece siempre que cualquier tejido resulta dañado y hace que el individuo reaccione apartando el estímulo doloroso. Incluso una actividad tan sencilla como estar sentado durante un período prolongado sobre el isquion puede provocar una destrucción tisular debido a la ausencia de flujo sanguíneo en la piel que quede comprimida por el peso del cuerpo. Cuando la piel comienza a doler a raíz de la isquemia, la persona normalmente cambia el apoyo del peso inconscientemente. Sin embargo, alguien que haya perdido la sensibilidad dolorosa, como sucede después de una lesión en la médula espinal, no llega a sentir este efecto y, por tanto, no se mueve. Esta situación pronto deriva en una excoriación y en la descamación total de la piel en las zonas de presión. Tipos de dolor y sus cualidades: dolor rápido y dolor lento El dolor se ha clasificado en dos tipos fundamentales: dolor rápido y dolor lento. El dolor rápido se siente en cuestión de 0,1 s después de haber aplicado el estímulo correspondiente, mientras que el dolor lento no empieza hasta pasado un mínimo de 1 s y a continuación crece con lentitud a lo largo de muchos segundos y en ocasiones hasta minutos. Durante el desarrollo de este capítulo veremos que son diferentes las vías de conducción para estos dos tipos de dolor y que cada una de ellas posee unas cualidades específicas. El dolor rápido también se describe con otros muchos nombres alternativos, como dolor intenso, dolor punzante, dolor agudo y dolor eléctrico. Este tipo de dolor se siente cuando se clava una aguja en la piel, cuando se corta con un cuchillo o cuando sufre una quemadura intensa. También se percibe cuando la piel se ve sometida a una sacudida eléctrica. El dolor rápido y agudo no se siente en los tejidos más profundos del organismo. El dolor lento también se designa con muchos nombres, entre ellos dolor lento urente, dolor sordo, dolor pulsátil, dolor nauseoso y dolor crónico. Este tipo de dolor suele ir asociado a una destrucción tisular. El dolor lento tiene la capacidad de propiciar un sufrimiento casi insoportable y prolongado. Puede darse en la piel y casi también en cualquier tejido u órgano profundo.
Receptores para el dolor y su estimulación Los receptores para el dolor son terminaciones nerviosas libres Los receptores para el dolor de la piel y de otros tejidos siempre son terminaciones nerviosas libres. Se encuentran extendidos por las capas superficiales de la piel así como en ciertos tejidos internos, como el periostio, las paredes arteriales, las superficies articulares, y las hoces y la tienda en la bóveda craneal. La mayoría de los demás tejidos profundos no reciben más que terminaciones dispersas para el dolor; no obstante, cualquier daño tisular generalizado puede acumularse hasta originar el tipo de dolor sordo, crónico y lento en la mayoría de estas zonas Tres tipos de estímulos excitan los receptores para el dolor: mecánicos, térmicos y químicos El dolor puede despertarse por múltiples tipos de estímulos, que se dividen en estímulos dolorosos mecánicos, térmicos y químicos. A grandes rasgos, el dolor rápido se suscita a partir de los tipos de estímulo mecánico y térmico, mientras que el dolor lento puede surgir con cualquiera de los tres. Algunos de los productos que excitan el dolor de tipo químico son bradicinina, serotonina, histamina, iones potasio, ácidos, acetilcolina y enzimas proteolíticas. Además, las prostaglandinas y la sustancia P favorecen la sensibilidad de las terminaciones para el dolor, pero no las activan directamente. Los compuestos químicos resultan especialmente importantes para estimular el tipo de dolor lento y molesto que ocurre después de una lesión tisular. Naturaleza no adaptativa de los receptores para el dolor Al revés que la mayoría de los otros receptores sensitivos del cuerpo, la adaptación de los receptores para el dolor es muy escasa y a veces nula en absoluto. En realidad, bajo ciertas condiciones, la excitación de las fibras para el dolor crece cada vez más, sobre todo en el tipo lento, sordo, nauseoso, si el estímulo doloroso persiste. Este aumento de la sensibilidad en los receptores para el dolor se llama hiperalgesia. Puede comprenderse con rapidez la importancia que tiene esta falta de adaptación, pues permite que el dolor mantenga informada a la persona de la existencia de un estímulo perjudicial para los tejidos mientras su origen siga presente.
Importancia especial de los estímulos dolorosos químicos durante la lesión tisular Los extractos de tejido dañado provocan un dolor intenso cuando se inyectan bajo la piel normal. En estos elementos pueden observarse la mayoría de los productos antes citados que excitan los receptores para el dolor de tipo químico. Una sustancia que parece más dolorosa que las demás es la bradicinina. Los investigadores han propuesto que este podría ser el agente con una mayor responsabilidad sobre el dolor generado después de un daño tisular. Asimismo, la intensidad dolorosa mantiene una relación con el incremento local en la concentración de los iones potasio o con la elevación de las enzimas proteolíticas que ataquen directamente las terminaciones nerviosas y despierten dolor al volver más permeables las membranas de los nervios a los iones. La isquemia tisular como causa de dolor Cuando queda bloqueado el flujo sanguíneo hacia un tejido, este suele volverse muy doloroso en cuestión de unos minutos. Cuanto mayor sea el metabolismo de este tejido, más rápida será la aparición del dolor. Por ejemplo, si se coloca un manguito para la presión arterial alrededor del brazo y se hincha hasta que cesa el flujo de sangre arterial, el ejercicio de los músculos del antebrazo a veces puede originar un dolor muscular en un plazo de 15 a 20 s. Si no se realiza esta acción, el dolor puede tardar de 3 a 4 min en surgir, aunque el flujo de sangre al músculo siga siendo nulo. Una de las causas propuestas para explicar el dolor que existe durante la isquemia es la acumulación de grandes cantidades de ácido láctico en los tejidos, surgido a raíz del metabolismo anaerobio (es decir, metabolismo sin oxígeno). También es probable que otros productos químicos, como la bradicinina y las enzimas proteolíticas, se formen en los tejidos debido al daño celular y que la presencia de estos agentes, sumada a la del ácido láctico, estimule las terminaciones nerviosas para el dolor. El espasmo muscular como causa de dolor El espasmo muscular también es una causa frecuente de dolor y representa el fundamento de muchos síndromes clínicos dolorosos. En este caso, probablemente obedece en parte al efecto directo que ejerce el espasmo muscular sobre la estimulación de los receptores para el dolor mecanosensibles, pero también podría desprenderse del efecto indirecto causado por este fenómeno al comprimir los vasos sanguíneos y generar una isquemia. Por añadidura, el espasmo acelera el metabolismo del tejido muscular, lo que acentúa aún más la isquemia relativa, y crea las condiciones ideales para la liberación de sustancias químicas inductoras de dolor.
Vías dobles para la transmisión de las señales de dolor en el sistema nervioso central A pesar de que todos los receptores para el dolor consisten en terminaciones nerviosas libres, estas estructuras utilizan dos vías distintas para transmitir sus señales respectivas hacia el sistema nervioso central. Ambas guardan una correspondencia básica con los dos tipos de dolor: una vía para el dolor rápido agudo y otra vía para el dolor lento crónico. Fibras periféricas para el dolor: fibras «rápidas» y «lentas» Las señales correspondientes al dolor rápido agudo nacen con estímulos dolorosos de tipo mecánico o térmico. Su transmisión sigue los nervios periféricos hasta la médula espinal a través de pequeñas fibras de tipo Aδ a una velocidad entre 6 y 30 m/s. Por el contrario, el dolor de carácter lento crónico se suscita sobre todo a partir de los estímulos químicos correspondientes, pero a veces también con estímulos mecánicos o térmicos persistentes. Este dolor lento crónico llega a la médula espinal por medio de las fibras de tipo C a una velocidad entre 0,5 y 2 m/s. Debido a este doble sistema de inervación para el dolor, un estímulo brusco de este carácter a menudo genera una sensación dolorosa «doble»: un dolor rápido agudo que llega al cerebro a través de la vía de las fibras Aδ, seguido más o menos 1 s después por un dolor lento que se transmite por la vía de las fibras C. El dolor agudo informa a gran velocidad sobre la situación lesiva y, por tanto, cumple una función importante para conseguir que la persona reaccione de inmediato y se aparte del estímulo. El dolor lento tiende a crecer con el tiempo. Esta sensación produce a la larga el dolor intolerable y obliga a que la persona trate de mitigar su causa.
Vías dobles para el dolor en la médula y en el tronco del encéfalo: los fascículos neoespinotalámico y paleoespinotalámico Al penetrar en la médula espinal, las señales de dolor toman dos caminos hacia el encéfalo, a través: 1) del fascículo neoespinotalámico 2) del fascículo paleoespinotalámico. Fascículo neoespinotalámico para el dolor rápido Las fibras rápidas para el dolor de tipo Aδ transmiten básicamente esta sensación en la modalidad térmica aguda y mecánica. Acaban sobre todo en la lámina I (lámina marginal) de las astas dorsales.
Capacidad del sistema nervioso para localizar el dolor rápido en el cuerpo El dolor de tipo rápido agudo puede localizarse con mucha mayor precisión en las diversas partes del cuerpo que el dolor lento crónico. Sin embargo, cuando no se estimulan más que los receptores para el dolor, sin una activación simultánea de los receptores táctiles, incluso el dolor rápido puede estar poco localizado, muchas veces solo con un margen de unos 10 cm alrededor de la zona en cuestión. Con todo, si la estimulación afecta a la vez a los receptores táctiles que activan el sistema de la columna dorsal-lemnisco medial, la localización puede ser prácticamente exacta. Glutamato, el neurotransmisor más probable de las fibras para el dolor rápido de tipo Aδ Se piensa que el glutamato es la sustancia neurotransmisora segregada en la médula espinal por las terminaciones de las fibras nerviosas para el dolor de tipo Aδ. El glutamato es uno de los transmisores excitadores que recibe un uso más generalizado en el sistema nervioso central, cuya acción normalmente no dura nada más que unos pocos milisegundos. Vía paleoespinotalámica para la transmisión del dolor lento crónico La vía paleoespinotalámica es un sistema mucho más antiguo y básicamente transmite el dolor procedente de las fibras periféricas de tipo C dotado de un carácter lento crónico, aunque también transporta algunas señales correspondientes a las fibras de tipo Aδ. En esta vía, dichas fibras periféricas acaban en la médula espinal casi en su integridad entre las láminas II y III de las astas dorsales, que en conjunto reciben el nombre de sustancia gelatinosa.
Sustancia P, el neurotransmisor más probable de las terminaciones nerviosas con un carácter lento crónico de tipo C Las investigaciones señalan que los terminales de las fibras para el dolor de tipo C que entran en la médula espinal segregan los transmisores glutamato y sustancia P. El glutamato actúa de manera inmediata y solo dura unos pocos milisegundos. La sustancia P se libera con mucha mayor lentitud, acumulándose su concentración durante un período de segundos o incluso de minutos. De hecho, se ha propuesto que la sensación dolorosa «doble» que se percibe después de un pinchazo podría derivar en parte de la circunstancia de que el glutamato proporciona el componente que le corresponde con mayor rapidez, mientras que el de la sustancia P llega más rezagado. Con independencia de los detalles aún sin conocer, parece claro que el glutamato es el neurotransmisor implicado más a fondo en enviar el dolor rápido hacia el sistema nervioso central, y que la sustancia P se ocupa del dolor lento crónico.
Capacidad muy escasa del sistema nervioso para localizar con precisión la fuente del dolor transmitido por la vía lenta crónica La localización del dolor transmitido a través de la vía paleoespinotalámica es imprecisa. Por ejemplo, el dolor lento crónico normalmente no se puede localizar más que en una zona amplia del cuerpo, como un brazo o una pierna, pero no en un punto específico de ese brazo o de esa pierna. Este fenómeno encaja con las conexiones difusas polisinápticas que posee esta vía. Así se explica por qué los pacientes muchas veces experimentan serios problemas para descubrir la fuente de algunos tipos crónicos de dolor. Función de la formación reticular, el tálamo y la corteza cerebral en la apreciación del dolor La resección íntegra de las áreas sensitivas somáticas de la corteza cerebral no evita la percepción del dolor. Por tanto, es probable que los impulsos dolorosos que penetran en la formación reticular del tronco del encéfalo, el tálamo y otros centros inferiores del encéfalo provoquen la percepción consciente de esta sensación. Esto no significa que la corteza cerebral no tenga nada que ver con su captación normal; la estimulación eléctrica de las áreas corticales somatosensitivas propicia la percepción de un dolor leve en el ser humano más o menos en el caso del 3% de los puntos elegidos. Sin embargo, se cree que esta estructura representa un papel de especial importancia en la interpretación de las cualidades del dolor, aunque su percepción pueda ser una función principalmente de los centros inferiores. Interrupción quirúrgica de las vías para el dolor Cuando una persona sufre un dolor intenso e incoercible (en ocasiones, como consecuencia de la diseminación rápida de un cáncer) es necesario aliviarlo. Para proporcionar alivio del dolor pueden cortarse las vías nerviosas para el dolor en cualquier punto entre varios posibles. Si esta sensación asienta en la parte inferior del cuerpo, una cordotomía en la región torácica de la médula espinal a menudo la mitiga durante unas semanas o meses. Para realizar una cordotomía, se efectúa una sección medular parcial en el lado contrario al dolor a lo largo de su cuadrante anterolateral para interrumpir la vía sensitiva anterolateral. La cordotomía no siempre tiene éxito para calmar el dolor, por dos razones. En primer lugar, muchas fibras que transportan esta sensación y proceden de la parte superior del cuerpo no cruzan hacia el lado opuesto de la médula espinal hasta después de haber llegado al encéfalo, por lo que la cordotomía no las divide. En segundo lugar, el dolor suele volver varios meses más tarde, en parte como consecuencia de la sensibilización experimentada por otras vías que en condiciones normales son demasiado tenues como para resultar eficaces (p. ej., las vías dispersas por el cordón dorsolateral). Otro método operatorio experimental para atenuar el dolor ha consistido en cauterizar las regiones específicas encargadas de esta función en los núcleos intralaminares del tálamo, lo que normalmente remedia los tipos de dolor que generan sufrimiento mientras deja intacta la apreciación personal del dolor «agudo», que representa un importante mecanismo protector. Sistema de supresión del dolor (ANALGESIA) en el encéfalo y en la médula espinal El grado con el que cada persona reacciona frente al dolor varía tremendamente. Esta variación obedece en parte a una propiedad que posee el encéfalo en sí mismo para suprimir la entrada de señales dolorosas al sistema nervioso mediante la activación de un mecanismo para controlar el dolor, llamado sistema de analgesia. (1) la región gris periacueductal y las áreas periventriculares del mesencéfalo y la parte superior de la protuberancia que rodean al acueducto de Silvio y a las porciones del tercer y del cuarto ventrículos. Desde estas zonas, las neuronas envían señales hacia (2) el núcleo magno del rafe, un núcleo delgado de la línea media situado en las partes inferior de la protuberancia y superior del bulbo raquídeo, y el núcleo reticular paragigantocelular, que ocupa una posición lateral en este último. A partir de estas estructuras, se transmiten señales descendentes de segundo orden por las columnas dorsolaterales de la médula espinal hacia (3) un complejo inhibidor del dolor localizado en las astas dorsales de la médula espinal. A este nivel, las señales analgésicas tienen la capacidad de bloquear el dolor antes de su transmisión hacia el encéfalo.
La estimulación eléctrica de la región gris periacueductal o del núcleo magno del rafe es capaz de suprimir muchas señales de dolor potentes que penetran a través de las raíces medulares dorsales. Asimismo, la activación de regiones que excitan la región gris periacueductal a niveles aún más altos del cerebro también puede suprimir el dolor. Entre ellas se cuentan: 1) los núcleos periventriculares del hipotálamo, que quedan adyacentes al tercer ventrículo 2) en menor medida, el fascículo prosencefálico medial, también en el hipotálamo. Diversas sustancias transmisoras participan en el sistema analgésico, especialmente la encefalina y la serotonina. La serotonina hace que las neuronas medulares locales liberen también encefalina. Se cree que la encefalina propicia una inhibición presináptica y postsináptica de las fibras para el dolor de tipo C y Aδ al hacer sinapsis en las astas dorsales. Por tanto, el sistema de analgesia es capaz de bloquear las señales de dolor en su punto de entrada inicial a la médula espinal.
Sistema de opioides cerebrales: endorfinas y encefalinas Hace más de 40 años se descubrió que la inyección de una cantidad minúscula de morfina en el núcleo periventricular que rodea al tercer ventrículo o en la región gris periacueductal del tronco del encéfalo provoca un grado extremo de analgesia. En los estudios posteriores se ha observado que los productos de tipo morfina, especialmente los opioides, actúan sobre otros muchos puntos del sistema de analgesia, entre ellos las astas dorsales de la médula espinal. Dado que la mayoría de las sustancias químicas que modifican la excitabilidad neuronal lo hacen actuando sobre los receptores sinápticos, se supuso que los «receptores de morfina» pertenecientes al sistema de analgesia deben estar destinados a algún neurotransmisor de tipo morfínico que posea una secreción de origen natural en el encéfalo. Por tanto, se emprendió una amplia búsqueda en pos del opioide natural encefálico. En la actualidad se han descubierto alrededor de una docena de tales sustancias opioides en diferentes puntos del sistema nervioso. Todos ellos son productos de degradación de tres grandes moléculas proteicas: proopiomelanocortina, proencefalina y prodinorfina. Entre los más importantes de estos compuestos figuran la β-endorfina, la metencefalina, la leuencefalina y la dinorfina. Las dos encefalinas están presentes en el tronco del encéfalo y en la médula espinal, dentro de las porciones del sistema de analgesia descritas antes, y la β-endorfina lo está en el hipotálamo y en la hipófisis. La dinorfina se encuentra básicamente en las mismas zonas que las encefalinas, pero en una cantidad mucho menor. Por tanto, aunque no se conocen por completo los detalles sobre el funcionamiento de los opioides cerebrales, la activación del sistema de analgesia por parte de las señales nerviosas que llegan a las regiones gris periacueductal y periventricular, o la inactivación de las vías para el dolor a cargo de los fármacos de tipo morfina, es capaz de suprimir casi en su integridad muchas de las señales dolorosas que entran a través de los nervios periféricos
Dolor referido Muchas veces una persona siente dolor en una parte del cuerpo situada bastante alejada del tejido que lo origina. Este fenómeno se llama dolor referido. Por ejemplo, su presencia en una de las vísceras a menudo queda remitida a una región de la superficie corporal. El conocimiento de sus diversos tipos resulta importante en el diagnóstico clínico porque en muchas dolencias viscerales el dolor referido es el único signo clínico.
Dolor visceral El dolor procedente de las diferentes vísceras del abdomen y del tórax es uno de los escasos criterios que pueden utilizarse para diagnosticar una inflamación visceral, las enfermedades infecciosas y otros padecimientos a este nivel. Muchas veces, las vísceras no poseen receptores sensitivos para ninguna otra modalidad de sensibilidad, salvo el dolor. Asimismo, el dolor visceral difiere del dolor superficial en varios aspectos importantes. Una de las discrepancias más importantes entre el dolor derivado de la superficie y el dolor visceral consiste en que los daños de tipo muy localizado en las vísceras rara vez originan un dolor intenso. Por ejemplo, un cirujano puede dividir el intestino del todo en dos partes con un paciente despierto sin causar un dolor apreciable. En cambio, cualquier fenómeno que produzca una estimulación difusa de las terminaciones nerviosas para el dolor en una víscera provoca un dolor intenso. Por ejemplo, la isquemia ocasionada por la oclusión del riego sanguíneo en una región intestinal amplia estimula muchas fibras difusas para el dolor al mismo tiempo y puede desembocar en un dolor terrible.
«Dolor parietal» provocado por enfermedades viscerales Cuando una enfermedad afecta a una víscera, el proceso patológico a menudo se propaga al peritoneo parietal, la pleura o el pericardio. Estas superficies parietales, lo mismo que la piel, reciben una amplia inervación dolorosa de los nervios raquídeos periféricos. Por tanto, el dolor procedente de la pared que cubre a una víscera con frecuencia tiene un carácter agudo. Un ejemplo puede subrayar la diferencia entre este dolor y el dolor visceral auténtico; una incisión con un bisturí a través del peritoneo parietal resulta muy dolorosa, mientras que un corte similar en el peritoneo visceral o en la pared intestinal no lo es tanto, o ni siquiera genera ningún dolor. Localización del dolor visceral: vías de transmisión del dolor «visceral» y «parietal» Por diversas razones, muchas veces cuesta localizar el dolor procedente de las diferentes vísceras. En primer lugar, el cerebro del paciente no posee experiencia de primera mano sobre la existencia de los distintos órganos internos; por tanto, cualquier dolor que tenga un origen interno no puede localizarse más que de un modo general. En segundo lugar, las sensaciones abdominales y torácicas se transmiten a través de dos vías hacia el sistema nervioso central: la vía visceral verdadera y la vía parietal. El dolor visceral verdadero recurre a las fibras sensitivas para el dolor contenidas en los haces de nervios autónomos, y las sensaciones resultan referidas a unas regiones de la superficie corporal muchas veces alejadas del órgano doloroso. Por el contrario, las sensaciones parietales se transportan directamente desde el peritoneo parietal, la pleura o el pericardio hacia los nervios raquídeos locales, y suelen quedar localizadas directamente sobre la zona dolorosa. En la médula espinal entre los segmentos C3 y T5. Por tanto, según se muestra en la figura 49-6, el dolor de origen cardíaco queda referido a la zona lateral del cuello, a la que cubre el hombro y los músculos pectorales, desciende por el brazo y llega al área subesternal de la parte alta del tórax. Estas son las zonas de la superficie corporal que envían sus propias fibras nerviosas somatosensitivas hacia los segmentos medulares C3 a T5. Lo más habitual es que el dolor asiente en el lado izquierdo en vez de en el derecho debido a que es mucho más frecuente la participación de este lado del corazón en una coronariopatía.
Cefalea Las cefaleas son un tipo de dolor referido a la superficie de la cabeza desde sus estructuras profundas. Algunas derivan de estímulos dolorosos que nacen en el interior del cráneo, pero otras lo hacen de un dolor cuyo origen está fuera, como en el caso de los senos nasales. Cefalea de origen intracraneal Zonas sensibles al dolor en la bóveda craneal Los tejidos cerebrales en sí son casi totalmente insensibles al dolor. Incluso el corte o la estimulación eléctrica de las áreas sensitivas de la corteza cerebral solo llega a generarlo en ocasiones; en cambio, provoca unos tipos de parestesias en forma de pinchazos sobre la región corporal representada en la porción de la corteza sensitiva estimulada. Así pues, es probable que el dolor correspondiente a la cefalea no esté causado en una gran proporción o en su mayoría por el daño del propio cerebro. Por el contrario, la tracción sobre los senos venosos que rodean el cerebro, la lesión de la tienda o el estiramiento de la duramadre en la base del encéfalo tienen la capacidad de despertar un dolor intenso que se identifica como una cefalea. Asimismo, casi cualquier clase de estímulo que traumatice, aplaste o estire los vasos sanguíneos de las meninges puede causar una cefalea. Una estructura especialmente sensible es la arteria meníngea media, y los neurocirujanos se preocupan de anestesiar este vaso específicamente cuando realizan una operación cerebral con uso de anestesia local. Zonas de la cabeza a las que queda referida una cefalea intracraneal La estimulación de los receptores para el dolor situados en la bóveda craneal por encima de la tienda, incluida la cara superior de esta última, desencadena impulsos dolorosos en la porción cerebral del quinto par y, por tanto, ocasiona una cefalea referida hacia la mitad anterior de la cabeza en las regiones superficiales inervadas por el componente somatosensitivo del quinto par craneal.
Tipos de cefalea intracraneal Cefalea de la meningitis Una de las cefaleas más intensas es la secundaria a la meningitis, proceso que causa una inflamación de todas las meninges, incluidas las áreas sensibles de la duramadre y las que rodean los senos venosos. Esta grave alteración puede provocar una cefalea extrema con dolor referido a toda la cabeza. Cefalea ocasionada por un descenso en la presión del líquido cefalorraquídeo La retirada de apenas 20 ml de líquido del conducto raquídeo, sobre todo si la persona permanece en posición vertical, muchas veces produce una intensa cefalea intracraneal. La extracción de una cantidad de líquido semejante elimina parte del componente de flotación cerebral que normalmente aporta el líquido cefalorraquídeo. El peso del cerebro estira y deforma las diversas superficies de la duramadre, y así desencadena el dolor que causa la cefalea. Jaqueca La jaqueca o cefalea migrañosa es un tipo especial de cefalea que puede derivar de fenómenos vasculares anormales, aunque no se conoce su mecanismo exacto. Las jaquecas suelen comenzar con diversas sensaciones prodrómicas, como náuseas, pérdida de la visión en parte del campo visual, auras visuales y otros tipos de alucinaciones sensitivas. En general, los síntomas prodrómicos empiezan de 30 min a 1 h antes de iniciarse la cefalea. Cualquier teoría capaz de explicar la jaqueca también ha de explicar estos síntomas prodrómicos. Una de las teorías sobre la causa de las jaquecas dice que una emoción o una tensión prolongada ocasionan un vasoespasmo reflejo de parte de las arterias de la cabeza, entre ellas las que irrigan el cerebro. En teoría, el vasoespasmo produce una isquemia de ciertas porciones cerebrales, que es la responsable de los síntomas prodrómicos. A continuación, como consecuencia de la isquemia intensa, algo sucede en las paredes vasculares, tal vez el agotamiento de la contracción del músculo liso, que condiciona un estado de flacidez e incapacidad para mantener el tono vascular durante 24 a 48 h. La presión arterial de los vasos hace que se dilaten y palpiten con intensidad, y se propone que el estiramiento excesivo de las paredes arteriales (incluidas algunas arterias extracraneales, como la temporal) despierta el dolor auténtico de las jaquecas. Otras teorías sobre la causa de las jaquecas hablan de la diseminación de la depresión cortical, de alteraciones psicológicas y de un vasoespasmo originado por el exceso local de potasio en el líquido extracelular cerebral. Puede existir una predisposición genética a las jaquecas, pues en el 65 al 90% de los casos se han recogido antecedentes familiares positivos. Las jaquecas también aparecen en las mujeres con una frecuencia aproximadamente doble que en los hombres. Cefalea alcohólica Como muchas personas saben por propia experiencia, la cefalea suele seguir a un consumo excesivo de alcohol. Es probable que el alcohol, debido a su naturaleza tóxica para los tejidos, irrite directamente las meninges y genere el dolor intracraneal. La deshidratación puede tener también parte de responsabilidad en la «resaca» posterior a un exceso alcohólico; la hidratación suele atenuar la cefalea y otros síntomas de la resaca, aunque no los elimina. Tipos extracraneales de cefalea Cefalea resultante de un espasmo muscular La tensión emocional a menudo hace que muchos de los músculos de la cabeza, sobre todo los que se insertan en el cuero cabelludo y la musculatura cervical que se fija en el occipucio, queden espásticos, y se propone que este mecanismo es una de las causas más frecuentes de cefalea. Cabe presumir que el dolor de los músculos espásticos de la cabeza quede referido a las zonas cefálicas que los cubren y genere el mismo tipo de cefalea que las lesiones intracraneales. Cefalea ocasionada por la irritación de las estructuras nasales y paranasales Las mucosas de la nariz y de los senos paranasales son sensibles al dolor, pero no tan intensamente. No obstante, una infección u otros procesos irritantes en extensas regiones de las estructuras nasales muchas veces se suman y propician una cefalea que queda referida detrás de los ojos o, en el caso de la infección del seno frontal, a las superficies frontales de la frente y del cuero cabelludo. Transmisión de señales térmicas en el sistema nervioso En general, las señales térmicas se transmiten por vías paralelas a las que siguen las señales dolorosas. Al entrar en la médula espinal, ascienden o descienden unos cuantos segmentos por el fascículo de Lissauer y después terminan sobre todo en las láminas I, II y III de las astas dorsales: las mismas que en el caso del dolor. Después de un cierto grado de procesamiento en una neurona medular o en más, las señales se incorporan a fibras térmicas ascendentes largas que cruzan hacia el fascículo sensitivo anterolateral opuesto y acaban en: 1) la formación reticular del tronco del encéfalo, y 2) el complejo ventrobasal del tálamo. Unas pocas señales térmicas también llegan a la corteza sensitiva somática del cerebro desde el complejo ventrobasal. A veces, mediante estudios con microelectrodos, se ha observado que una neurona del área sensitiva somática cortical I es sensible directamente a los estímulos de frío o de calor en una zona específica de la piel. Sin embargo, en el ser humano, la eliminación de la circunvolución poscentral de la corteza en su integridad va a reducir la capacidad de distinguir gradaciones de temperatura, pero no a abolirla.
Nuestras percepciones de las señales del cuerpo y del mundo que nos rodea están mediadas por un complejo sistema de receptores sensitivos que detectan estímulos como el tacto, el sonido, la luz, el dolor, el frío y el calor. El propósito del presente capítulo consiste en exponer los mecanismos básicos por los que estos receptores transforman los estímulos sensitivos en señales nerviosas que a continuación son enviadas y procesadas en el sistema nervioso central.
Sensibilidad diferencial de los receptores ¿Cómo dos tipos distintos de receptores sensitivos detectan clases diferentes de estímulos sensitivos? La respuesta es la siguiente: por sus sensibilidades diferenciales. Es decir, cada tipo de receptor resulta muy sensible a una clase de estímulo sensitivo para el que está diseñado y en cambio es casi insensible a otras clases. De este modo, los conos y los bastones de los ojos son muy sensibles a la luz, pero casi totalmente insensibles a una situación de calor, frío, presión sobre los globos oculares o cambios químicos en la sangre dentro de los límites normales. Los osmorreceptores de los núcleos supraópticos en el hipotálamo detectan variaciones minúsculas en la osmolalidad de los líquidos corporales, pero nunca se ha visto que respondan al sonido. Por último, los receptores cutáneos para el dolor casi nunca se estimulan con los estímulos corrientes de tacto o de presión, pero pasan a estar muy activos en el momento en que adquieren la intensidad suficiente para dañar a los tejidos.
Modalidad sensitiva: el principio de la «línea marcada» Cada uno de los principales tipos sensitivos que podemos experimentar, dolor, tacto, visión, sonido, etc., se llama modalidad de sensación. Con todo, pese al hecho de que nosotros percibimos estas diversas modalidades, las fibras nerviosas únicamente transmiten impulsos. Por tanto, ¿cómo es que distintas fibras nerviosas transmiten modalidades diferentes de sensación? La respuesta señala que cada fascículo nervioso termina en un punto específico del sistema nervioso central y el tipo de sensación vivida cuando se estimula una fibra nerviosa queda determinado por la zona del sistema nervioso a la que conduce esta fibra. Por ejemplo, si se estimula una fibra para el dolor, la persona percibe esta sensación sea cual sea el tipo de estímulo que la excite. Puede ser la electricidad, el recalentamiento de la fibra, su aplastamiento o la activación de la terminación nerviosa para el dolor cuando las células tisulares sufren una lesión. En todos estos casos, la persona percibe dolor. Análogamente, si se estimula una fibra táctil por la excitación eléctrica de un receptor para el tacto o por cualquier otro mecanismo, la persona percibe sensación de tacto porque dichas fibras conducen hasta las áreas específicas del tacto en el cerebro. En este mismo sentido, las fibras procedentes de la retina ocular terminan en las áreas visuales del cerebro, las del oído acaban en las áreas auditivas y las térmicas en las áreas para la temperatura. Esta especificidad de las fibras nerviosas para transmitir nada más que una modalidad de sensación se llama principio de la línea marcada.
Transducción de estímulos sensitivos en impulsos nerviosos Corrientes eléctricas locales en las terminaciones nerviosas: potenciales de receptor Todos los receptores sensitivos tienen un rasgo en común. Cualquiera que sea el tipo de estímulo que les excite, su efecto inmediato consiste en modificar su potencial eléctrico de membrana. Este cambio en el potencial se llama potencial de receptor. Mecanismos de los potenciales de receptor Los diversos receptores pueden excitarse siguiendo alguno de los siguientes modos de generar potenciales de receptor: 1) por deformación mecánica del receptor, que estire su membrana y abra los canales iónicos 2) por la aplicación de un producto químico a la membrana, que también abra los canales iónicos 3) por un cambio de la temperatura de la membrana, que modifique su permeabilidad 4) por los efectos de la radiación electromagnética, como la luz que incide sobre un receptor visual de la retina, al modificar directa o indirectamente las características de la membrana del receptor y permitir el flujo de iones a través de sus canales. Estos cuatro medios de excitar a los receptores guardan una correspondencia general con los diferentes tipos de receptores sensitivos conocidos. En todos los casos, la causa básica del cambio en el potencial de membrana es una modificación en la permeabilidad de la membrana del receptor, que permite la difusión iónica con mayor o menor facilidad a través de la membrana y variar así el potencial transmembrana. Amplitud del potencial de receptor máximo La amplitud máxima de la mayoría de los potenciales de receptor sensitivos es de unos 100 mV, pero este valor no se alcanza más que cuando la intensidad del estímulo correspondiente es altísima. Más o menos se trata del mismo voltaje máximo registrado en los potenciales de acción y también es el cambio que sucede cuando la membrana adquiere una permeabilidad máxima a los iones sodio. Relación del potencial de receptor con los potenciales de acción Cuando el potencial de receptor sube por encima del umbral necesario para desencadenar potenciales de acción en la fibra nerviosa adscrita al receptor, se produce su aparición.
Mecanismo de adaptación de los receptores El mecanismo de adaptación varía con cada tipo de receptor, básicamente lo mismo que la producción de un potencial de receptor constituye una propiedad individual. Por ejemplo, en el ojo, los conos y los bastones se adaptan al modificarse las concentraciones de sus sustancias químicas sensibles a la luz . En el caso de los mecanorreceptores, el que se ha estudiado con mayor detalle es el corpúsculo de Pacini. En este caso, la adaptación sucede de dos maneras. En primer lugar, el corpúsculo de Pacini es una estructura viscoelástica, por lo que si se aplica de repente una fuerza deformadora sobre uno de sus lados, esta fuerza se transmite al instante directamente al mismo lado de la fibra nerviosa central debido al componente viscoso del corpúsculo, lo que desencadena un potencial de receptor. Sin embargo, en unas pocas centésimas de segundo, el líquido contenido en su interior se redistribuye, de manera que deja de generarse el potencial de receptor. Así pues, este último surge al comienzo de la compresión pero desaparece en cuestión de una pequeña fracción de segundo, aunque siga presente su acción. El segundo mecanismo de adaptación del corpúsculo de Pacini, mucho más lento en su desarrollo, deriva de un proceso llamado acomodación, que sucede en la propia fibra nerviosa. Esto es, aunque por casualidad la fibra del núcleo central continúe deformada, el extremo de la fibra nerviosa se «acomoda» paulatinamente al estímulo. Esto tal vez obedezca a una «inactivación» progresiva de los canales de sodio en su membrana, lo que significa que el flujo de la corriente de este ion a través suyo hace que se cierren poco a poco, efecto que parece ocurrir en todos o en la mayoría de los canales de sodio de la membrana celular.
Los receptores de adaptación lenta detectan la intensidad continua del estímulo: los receptores «tónicos» Los receptores de adaptación lenta siguen transmitiendo impulsos hacia el cerebro mientras siga presente el estímulo (o al menos durante muchos minutos u horas). Por tanto, mantienen al cerebro constantemente informado sobre la situación del cuerpo y su relación con el medio. Por ejemplo, los impulsos procedentes de los husos musculares y de los aparatos tendinosos de Golgi ponen al sistema nervioso en condiciones de conocer el estado de contracción muscular y la carga soportada por el tendón muscular en cada instante. Otros receptores de adaptación lenta son los siguientes: 1) los pertenecientes a la mácula en el aparato vestibular; 2) los receptores para el dolor; 3) los barorreceptores del árbol arterial, y 4) los quimiorreceptores de los cuerpos carotídeo y aórtico. Debido a su capacidad para seguir transmitiendo información durante muchas horas, o incluso días, también se les denomina receptores tónicos. Los receptores de adaptación rápida detectan cambios en la intensidad del estímulo: «receptores de velocidad», «receptores de movimiento» o «receptores fásicos» Los receptores que se adaptan con rapidez no pueden utilizarse para transmitir una señal continua debido a que solo se activan cuando cambia la intensidad del estímulo. Con todo, reaccionan potentemente siempre que esté teniendo lugar un cambio de hecho. Por tanto, se llaman receptores de velocidad, receptores de movimiento o receptores fásicos. Así pues, en el caso del corpúsculo de Pacini, la presión brusca aplicada al tejido lo excita durante unos pocos milisegundos, y a continuación se acaba su excitación, aun cuando siga actuando. Sin embargo, más tarde transmite de nuevo una señal si se alivia esta presión. Es decir, el corpúsculo de Pacini resulta sumamente importante para comunicar al sistema nervioso las deformaciones rápidas de un tejido, pero no sirve de nada para transmitir información acerca de una situación constante en el organismo. Función predictiva de los receptores de velocidad Si se conoce la velocidad a la que tiene lugar un cambio en la situación corporal, se podrá predecir cuál será el estado del organismo a su juicio unos cuantos segundos o incluso minutos más tarde. Por ejemplo, los receptores existentes en los conductos semicirculares del aparato vestibular del oído detectan la velocidad a la que empieza a girar la cabeza cuando se toma una curva. Con esta información, una persona es capaz de pronosticar el grado de giro durante los 2 s siguientes y corregir el movimiento de las piernas por anticipado para no perder el equilibrio. En este mismo sentido, los receptores situados en las articulaciones o en su proximidad sirven para detectar las velocidades de movimiento que llevan las diferentes partes del cuerpo. Por ejemplo, cuando se corre, la información procedente de los receptores de velocidad articulares le permite al sistema nervioso vaticinar dónde estarán los pies en cualquier fracción exacta del próximo segundo. Por tanto, pueden transmitirse las señales motoras correspondientes a los músculos de las piernas para realizar todas las correcciones de su posición pertinentes como medida preventiva con el fin de no caerse. La pérdida de esta función predictiva impide correr a una persona.
CLASIFICACION RAPIDA DE LAS FIBRAS NERVIOSAS.
En la clasificación general, las fibras se dividen en los tipos A y C, y las de tipo A se subdividen aún en las fibras α, β, γ y δ. Las de tipo A son las típicas fibras mielínicas de tamaño grande y medio pertenecientes a los nervios raquídeos. Las de tipo C son las fibras nerviosas pequeñas amielínicas que conducen los impulsos a velocidades bajas. Estas últimas representan más de la mitad de las fibras sensitivas en la mayoría de los nervios periféricos, así como todas las fibras autónomas posganglionares. Obsérvese que unas cuantas fibras mielínicas grandes son capaces de transmitir impulsos a velocidades superiores a los 120 m/s, es decir, recorrer en 1 s una distancia que supera la longitud de un campo de fútbol. Por el contrario, las fibras más pequeñas transmiten impulsos incluso nada más que a 0,5 m/s, lo que supone 2 s para ir desde el dedo gordo del pie hasta la médula espinal. Clasificación alternativa empleada por los fisiólogos de la sensibilidad Ciertas técnicas de registro han permitido dividir las fibras de tipo Aα en dos subgrupos; no obstante, estos mismos métodos no son capaces de distinguir con facilidad entre las fibras Aβ y Aγ. Por tanto, los fisiólogos de la sensibilidad emplean a menudo la siguiente clasificación: Grupo Ia Fibras procedentes de las terminaciones anuloespirales de los husos musculares (con un diámetro medio de unos 17 μm; son fibras A de tipo α según la clasificación general). Grupo Ib Fibras procedentes de los órganos tendinosos de Golgi (con un diámetro medio de unos 16 μm; también son fibras A de tipo α). Grupo II Fibras procedentes de la mayoría de los receptores táctiles cutáneos aislados y de las terminaciones en ramillete de los husos musculares (con un diámetro medio de unos 8 μm; son fibras A de tipo β y γ según la clasificación general). Grupo III Fibras que transportan la temperatura, el tacto grosero y las sensaciones de dolor y escozor (con un diámetro medio de unos 3 μm; son fibras A de tipo δ según la clasificación general). Grupo IV Fibras amielínicas que transportan las sensaciones de dolor, picor, temperatura y tacto grosero (con un diámetro de 0,5 a 2 μm; son fibras de tipo C según la clasificación general).
Transmisión de señales de diferente intensidad por los fascículos nerviosos: sumación espacial y temporal Una de las características de toda señal que siempre ha de transportarse es su intensidad: por ejemplo, la intensidad del dolor. Los diversos grados de esta variable pueden transmitirse mediante un número creciente de fibras paralelas o enviando más potenciales de acción a lo largo de una sola fibra. Estos dos mecanismos se llaman, respectivamente, sumación espacial y sumación temporal. SUMACION ESPACIAL. Cada una de estas fibras se ramifica en cientos de minúsculas terminaciones nerviosas libres que sirven como receptores para el dolor. Todo el conglomerado formado por las fibras que proceden de una sola con frecuencia cubre una zona de piel cuyo diámetro llega a medir 5 cm. Este área se llama campo receptor de la fibra. El número de terminaciones es grande en su centro, pero disminuye hacia la periferia. En la figura también se puede observar que las fibrillas ramificadas se superponen con las derivadas de otras fibras para el dolor. Por tanto, un pinchazo en la piel suele estimular las terminaciones de muchas diferentes a la vez. Cuando el pinchazo ocurre en el centro del campo receptor de una fibra para el dolor concreta, su grado de estimulación es muy superior a si sucede en la periferia, porque el número de terminaciones nerviosas libres es mucho mayor en dicho caso. Sumación temporal Un segundo medio para transmitir señales de intensidad creciente consiste en acelerar la frecuencia de los impulsos nerviosos que recorren cada fibra, lo que se denomina sumación temporal.
Transmisión y procesamiento de las señales en grupos neuronales El sistema nervioso central está integrado por miles de millones de grupos neuronales; algunos de estos grupos contienen unas cuantas neuronas, mientras que otros presentan una cantidad enorme. Por ejemplo, toda la corteza cerebral podría considerarse un solo gran grupo neuronal. Otros casos similares los ofrecen los diversos componentes de los ganglios basales y los núcleos específicos del tálamo, el cerebelo, el mesencéfalo, la protuberancia y el bulbo raquídeo. Asimismo, toda la sustancia gris dorsal de la médula espinal podría tomarse como un gran grupo de neuronas. Cada grupo neuronal posee su propia organización especial que le hace procesar las señales de un modo particular y singular, lo que permite que el agregado total de grupos cumpla la multitud de funciones del sistema ner
Divergencia de las señales que atraviesan los grupos neuronales Muchas veces es importante que las señales débiles que penetran en un grupo neuronal acaben excitando a una cantidad mucho mayor de las fibras nerviosas que lo abandonan. Este fenómeno se llama divergencia. Existen dos tipos fundamentales de divergencia que cumplen unos propósitos totalmente diferentes. Esta divergencia significa sencillamente que una señal de entrada se disemina sobre un número creciente de neuronas a medida que atraviesa sucesivos órdenes de células en su camino. Es el tipo de divergencia característico de la vía corticoespinal en su labor de control sobre los músculos esqueléticos, en la que cada célula piramidal grande de la corteza motora es capaz de excitar hasta 10.000 fibras musculares cuando se halla en unas condiciones muy facilitadas.
Circuito neuronal con señales de salida excitadoras e inhibidoras En ocasiones, una señal de entrada en un grupo neuronal hace que una señal excitadora de salida siga una dirección y a la vez otra señal inhibidora vaya hacia otro lugar. Por ejemplo, al mismo tiempo que una señal excitadora se transmite a lo largo de una serie de neuronas en la médula espinal para provocar el movimiento hacia delante de una pierna, otra señal inhibidora viaja a través de una colección distinta de neuronas para inhibir los músculos de la parte posterior de la pierna a fin de que no se opongan al movimiento hacia delante. Este tipo de circuito es característico en el control de todos los pares de músculos antagonistas y se llama circuito de inhibición recíproca. Prolongación de una señal por un grupo neuronal: «posdescarga» Hasta ahora hemos considerado las señales que se transmiten simplemente a través de grupos neuronales. Sin embargo, en muchos casos, una señal que penetra en un grupo suscita una descarga de salida prolongada, llamada posdescarga, cuya duración va desde unos pocos milisegundos hasta muchos minutos después de que haya acabado la señal de entrada. Los mecanismos más importantes por los que sucede la posdescarga se describen en los apartados siguientes. Posdescarga sináptica Cuando las sinapsis excitadoras descargan sobre la superficie de las dendritas o del soma en una neurona, surge en ella un potencial eléctrico postsináptico que dura muchos milisegundos, especialmente cuando interviene alguna de las sustancias transmisoras sinápticas de acción prolongada. Circuito reverberante (oscilatorio) como causa de la prolongación de la señal Uno de los circuitos más importantes del sistema nervioso es el circuito reverberante u oscilatorio. Está ocasionado por una retroalimentación positiva dentro del circuito neuronal que ejerce una retroalimentación encargada de reexcitar la entrada del mismo circuito.
Emisión de señales continuas desde algunos circuitos neuronales Algunos circuitos neuronales emiten señales de salida de forma continua, incluso sin señales de entrada excitadoras. Al menos dos mecanismos pueden ocasionar este efecto: 1) la descarga neuronal intrínseca continua, y 2) las señales reverberantes continuas. Descarga continua ocasionada por la excitabilidad neuronal intrínseca Las neuronas, igual que otros tejidos excitables, descargan de forma repetida si el nivel del potencial de membrana excitador sube por encima de un cierto valor umbral. Los potenciales de membrana de muchas neuronas son suficientemente altos incluso en situaciones normales como para hacer que emitan impulsos de forma continua. Este fenómeno sucede sobre todo en gran parte de las neuronas del cerebelo, lo mismo que en la mayoría de las interneuronas de la médula espinal. Las frecuencias de emisión de impulsos en estas células pueden aumentar si reciben señales excitadoras o disminuir si son inhibidoras; estas últimas a menudo tienen la capacidad de rebajar la frecuencia de disparo hasta cero. Señales continuas emitidas desde circuitos reverberantes como medio para transmitir información Un circuito reverberante que no alcance un grado de fatiga suficiente como para detener la reverberación es una fuente de impulsos continuos. Además, los impulsos excitadores que penetran en el grupo reverberante pueden incrementar la señal de salida, mientras que la inhibición puede reducir o incluso extinguir la señal. Emisión de señales rítmicas
Muchos circuitos neuronales emiten señales de salida rítmicas: por ejemplo, una señal respiratoria rítmica nace en los centros respiratorios del bulbo raquídeo y de la protuberancia. La señal rítmica respiratoria se mantiene de por vida. Otras señales rítmicas, como las que causan los movimientos de rascado de un perro con la pata trasera o las actividades motoras de cualquier animal al caminar, requieren la existencia de estímulos de entrada en los respectivos circuitos para desencadenar las señales rítmicas. Se ha observado que todas o casi todas las señales rítmicas estudiadas por medios experimentales derivan de circuitos reverberantes o de una sucesión suya en serie que suministra señales excitadoras o inhibidoras de forma circular desde un grupo neuronal al siguiente.
Inestabilidad y estabilidad de los circuitos neuronales Casi cualquier parte del cerebro posee conexiones directas o indirectas con cualquier otra parte, lo que crea un serio problema. Si la primera porción excita a la segunda, la segunda a la tercera, la tercera a la cuarta y así sucesivamente hasta que al final la señal reexcite la primera porción, está claro que una señal excitadora que penetre a cualquier nivel del cerebro detonaría un ciclo continuo de reexcitaciones por todas partes. Si este ciclo llegara a producirse, el cerebro quedaría inundado por una nube de señales reverberantes sin control alguno, señales que no estarían portando ninguna información pero que, no obstante, sí estarían ocupando los circuitos cerebrales de manera que sería imposible transmitir cualquier señal realmente informativa. Tal efecto acontece en amplias regiones cerebrales durante las convulsiones epilépticas. ¿Cómo evita el sistema nervioso central que suceda este efecto permanentemente? La respuesta reside fundamentalmente en dos mecanismos básicos que funcionan a lo largo de todo el sistema nervioso central: 1) los circuitos inhibidores, y 2) la fatiga de las sinapsis. Circuitos inhibidores como mecanismo para estabilizar la función del sistema nervioso Dos tipos de circuitos inhibidores sirven para impedir la difusión excesiva de las señales por extensas regiones del encéfalo: 1) los circuitos de retroalimentación inhibidores que vuelven desde el extremo terminal de una vía hacia las neuronas excitadoras iniciales de esa misma vía (estos circuitos existen prácticamente en todas las vías nerviosas sensitivas e inhiben tanto sus neuronas de entrada como las neuronas intermedias cuando el extremo terminal está demasiado excitado), y 2) ciertos grupos neuronales que ejercen un control inhibidor global sobre regiones generalizadas del cerebro (p. ej., gran parte de los ganglios basales ejercen influencias inhibidoras sobre todo el sistema de control muscular).
Corrección automática a corto plazo de la sensibilidad de la vía mediante el mecanismo de la fatiga A continuación vamos a aplicar este fenómeno de la fatiga a otras vías cerebrales. Cuando están sometidas a un uso excesivo, suelen acabar fatigándose, por lo que desciende su sensibilidad. A la inversa, las que están infrautilizadas se encuentran descansadas y sus sensibilidades aumentan. Por tanto, la fatiga y su recuperación constituyen un medio importante a corto plazo para moderar la sensibilidad de los diferentes circuitos del sistema nervioso. Estas funciones sirven para mantener los circuitos en operación dentro de unos márgenes de sensibilidad que permitan su funcionamiento eficaz. Cambios a largo plazo en la sensibilidad sináptica ocasionados por la regulación al alza o a la baja de los receptores sinápticos La sensibilidad a largo plazo de las sinapsis puede cambiar tremendamente si la cantidad de proteínas receptoras presentes en los puntos sinápticos se regula al alza en una situación de baja actividad, y a la baja cuando haya una hiperactividad. El mecanismo de este proceso es el siguiente: las proteínas receptoras están formándose constantemente en el sistema integrado por el retículo endoplásmico y el aparato de Golgi y son introducidas de modo continuo en la membrana sináptica de la neurona receptora. Sin embargo, cuando las sinapsis se emplean demasiado de manera que una cantidad excesiva de sustancia transmisora se combina con las proteínas receptoras, muchos de estos receptores quedan inactivados y retirados de la membrana sináptica. En efecto, es una suerte que la regulación al alza y a la baja de los receptores, lo mismo que otros mecanismos de control destinados a adecuar la sensibilidad sináptica, corrijan permanentemente esta propiedad en cada circuito hasta el nivel casi exacto necesario para su correcto funcionamiento. Piense por un momento en la seriedad de la situación nada más con que la sensibilidad de unos cuantos de estos circuitos fuera anormalmente alta; entonces cabría esperar la presencia casi continua de calambres musculares, convulsiones, alteraciones psicóticas, alucinaciones, tensión mental u otros trastornos nerviosos. Afortunadamente, los controles automáticos suelen reajustar la sensibilidad de los circuitos de vuelta dentro de unos límites de reactividad controlables en cualquier momento en que empiecen a estar demasiado activos o demasiado deprimidos.
El sistema nervioso carece de parangón en cuanto a la enorme complejidad de los procesos de pensamiento y acciones de control que es capaz de realizar. Cada minuto son literalmente millones los fragmentos de información que recibe procedentes de los distintos nervios y órganos sensitivos y a continuación integra todo este cúmulo para generar las respuestas que vaya a emitir el organismo.
DISEÑO GENERAL DEL SISTEMA NERVIOSO. «La neurona: unidad funcional básica del sistema nervioso central.» El sistema nervioso central contiene más de 100.000 millones de neuronas. Según los diversos tipos de neuronas, las conexiones sinápticas procedentes de las fibras aferentes pueden ser tan solo unos cientos o llegar hasta 200.000. Por el contrario, la señal de salida viaja por el único axón que abandona la neurona La mayoría de las actividades del sistema nervioso se ponen en marcha cuando las experiencias sensitivas excitan los receptores sensitivos, ya sean de carácter visual en los ojos, auditivo en los oídos, táctil en la superficie del organismo o de otros tipos. Estas experiencias sensitivas pueden desencadenar reacciones inmediatas del encéfalo, o almacenarse su recuerdo durante minutos, semanas o años y determinar reacciones corporales en algún momento futuro.
Esta información penetra en el sistema nervioso central a través de los nervios periféricos y se transporta de inmediato hasta múltiples zonas sensitivas en: 1) la médula espinal a todos sus niveles; 2) la formación reticular del bulbo raquídeo, la protuberancia y el mesencéfalo en el encéfalo; 3) el cerebelo; 4) el tálamo, y 5) áreas de la corteza cerebral.
«PROPORCIÓN MOTORA DEL SISTEMA NERVIOSO: EFECTORES.» El sistema nervioso regula diversas actividades del organismo, para eso, debe aprender a desempeñar las siguientes tareas: 1) la contracción de los músculos esqueléticos adecuados en todo el cuerpo 2) la contracción de la musculatura lisa de las vísceras, 3) la secreción de sustancias químicas activas por parte de las glándulas exocrinas y endocrinas en muchas zonas del organismo. En conjunto, estas actividades se denominan funciones motoras del sistema nervioso y los músculos y las glándulas reciben el nombre de efectores porque representan las estructuras anatómicas reales que ejecutan las funciones dictadas por las señales nerviosas. Un segundo elemento, llamado sistema nervioso autónomo, opera de forma paralela a su acción, estando encargado de controlar la musculatura lisa, las glándulas y otros sistemas corporales internos.
«FUNCIÓN INTEGRADORA» Una de las funciones más importantes del sistema nervioso consiste en elaborar la información que le llega de tal modo que dé lugar a las respuestas motoras y mentales adecuadas. El encéfalo descarta más del 99% de toda la información sensitiva que recibe por carecer de interés o de importancia. Por ejemplo, corrientemente una persona no tiene conciencia de las diversas porciones de su cuerpo que están en contacto con la ropa, ni tampoco de la presión originada por el asiento sobre el que descansa. En este mismo sentido, solo llama la atención un objeto que ocupe el campo visual esporádicamente, e incluso los sonidos constantes de nuestro entorno suelen quedar relegados al inconsciente, pero cuando una información sensitiva importante excita la mente, de inmediato resulta encauzada hacia las regiones motoras e integradoras oportunas del encéfalo para suscitar las respuestas deseadas. Esta canalización y tratamiento de la información se denomina función integradora del sistema nervioso. Así, si una persona pone la mano sobre una estufa caliente, la respuesta instantánea pretendida consiste en levantarla. Esto deja paso a otras respuestas asociadas, como apartar todo el cuerpo de la estufa y a veces incluso gritar de dolor.
«ALMACENAMIENTO DE INFORMACIÓN: MEMORIA.»
La acumulación de la información es el proceso que llamamos memoria, y también constituye una función de las sinapsis. Cada vez que determinados tipos de señales sensitivas atraviesan una secuencia de sinapsis, estas adquieren una mayor capacidad para transmitir ese mismo tipo de señal la próxima vez, situación que llamamos facilitación. Después de que las señales sensitivas hayan recorrido las sinapsis en multitud de ocasiones, su facilitación es tan profunda que las señales generadas dentro del propio encéfalo también pueden originar la transmisión de impulsos a lo largo de la misma serie de sinapsis, incluso cuando no haya sido estimulada su entrada sensitiva. Este proceso otorga a la persona una percepción de estar experimentando sensaciones originales, aunque únicamente se trate de recuerdos. Una vez que los recuerdos están guardados en el sistema nervioso, pasan a formar parte de los mecanismos de procesamiento cerebral para el «pensamiento» en el futuro. Es decir, los procedimientos de deliberación del encéfalo comparan las experiencias sensitivas nuevas con los recuerdos acumulados; a continuación, estos últimos sirven para seleccionar la información sensitiva nueva que resulte más importante y encauzarla hacia las regiones correspondientes para el almacenamiento de la memoria a fin de permitir su uso en el futuro o hacia las regiones motoras para dar lugar a las respuestas corporales inmediatas.
«NIVELES DE FUNCIÓN DEL SISTEMA NERVIOSO.» El sistema nervioso humano ha heredado unas capacidades funcionales especiales correspondientes a cada etapa recorrida por el desarrollo evolutivo del hombre. A partir de este bagaje, los principales niveles del sistema nervioso central que presentan unas características funcionales específicas son tres: 1) el nivel medular: . Esta suposición dista mucho de la verdad. Incluso después de haber seccionado la médula espinal en la región cervical alta, seguirán ocurriendo muchas funciones medulares dotadas de una gran organización. Por ejemplo, los circuitos neuronales de la médula pueden originar: 1) los movimientos de la marcha 2) reflejos para retirar una parte del organismo de los objetos dolorosos 3) reflejos para poner rígidas las piernas para sostener el tronco en contra de la gravedad 4) reflejos que controlan los vasos sanguíneos locales, los movimientos digestivos o la excreción urinaria. En realidad, los niveles superiores del sistema nervioso no suelen operar enviando señales directamente hacia la periferia del cuerpo sino hacia los centros de control en la médula, simplemente «ordenando» que estos centros ejecuten sus funciones. 2) el nivel encefálico inferior o subcortical: actividades inconscientes del organismo están controladas por las regiones inferiores del encéfalo, es decir, el bulbo raquídeo, la protuberancia, el mesencéfalo, el hipotálamo, el tálamo, el cerebelo y los ganglios basales. El control del equilibrio constituye una función combinada entre las porciones más antiguas del cerebelo y la formación reticular del bulbo raquídeo, la protuberancia y el mesencéfalo. Los reflejos de la alimentación, como la salivación y el humedecimiento de los labios en respuesta al sabor de la comida, están regulados por regiones del bulbo raquídeo, la protuberancia, el mesencéfalo, la amígdala y el hipotálamo. Además, numerosos patrones emocionales, como la ira, la excitación, las respuestas sexuales, las reacciones al dolor y al placer, aún pueden darse una vez destruida gran parte de la corteza cerebral. 3) el nivel encefálico superior o cortical: la corteza cerebral resulta fundamental para la mayoría de los procesos de nuestro pensamiento, pero no puede funcionar por su cuenta. En realidad, son los centros encefálicos inferiores, y no la corteza, los que despiertan en ella la vigilia, abriendo así su banco de recuerdos a la maquinaria cerebral del razonamiento, cada porción del sistema nervioso cumple unas funciones específicas, pero es la corteza la que destapa todo un mundo de información almacenada para que la mente la use.
«SINAPSIS Y TIPOS DE SINAPSIS (QUÍMICAS O ELÉCTRICAS)» La información recorre el sistema nervioso central sobre todo bajo la forma de potenciales de acción nerviosos, llamados simplemente impulsos nerviosos, a través de una sucesión de neuronas, una después de la otra, existen 2 tipos de sinapsis: QUIMICA Y ELECTRICA. La mayoría de las sinapsis utilizadas para la transmisión de señales en el sistema nervioso central del ser humano son sinapsis químicas. En estas sinapsis, la primera neurona segrega un producto químico denominado neurotransmisor (a menudo llamado sustancia transmisora) a nivel de la terminación nerviosa, que a su vez actúa sobre las proteínas receptoras presentes en la membrana de la neurona siguiente para excitarla, inhibirla o modificar su sensibilidad de algún otro modo. Hasta hoy se han descubierto más de 40 neurotransmisores importantes. Entre las mejor conocidas figuran las siguientes: acetilcolina, noradrenalina, adrenalina, histamina, ácido γ-aminobutírico (GABA), glicina, serotonina y glutamato. En las sinapsis eléctricas los citoplasmas de las células adyacentes están conectados directamente por grupos de canales de iones llamados uniones en hendidura que permiten el movimiento libre de los iones desde el interior de una célula hasta el interior de la siguiente.
Aunque la mayoría de las sinapsis en el encéfalo son químicas, en el sistema nervioso central pueden coexistir sinapsis eléctricas y químicas. La transmisión bidireccional de las sinapsis eléctricas les permite colaborar en la coordinación de las actividades de grandes grupos de neuronas interconectadas.
Conducción «unidireccional» en las sinapsis químicas. Las sinapsis químicas poseen una característica sumamente importante que las convierte en un elemento muy conveniente para transmitir la mayor parte de las señales en el sistema nervioso. Esta característica hace posible que siempre conduzcan las señales en un solo sentido: es decir, desde la neurona que segrega el neurotransmisor, denominada neurona presináptica, hasta la neurona sobre la que actúa el transmisor, llamada neurona postsináptica. Este fenómeno es el principio de la conducción unidireccional de las sinapsis químicas y se aleja bastante de la conducción a través de las sinapsis eléctricas, que muchas veces transmiten señales en ambos sentidos. Un mecanismo de conducción unidireccional da la oportunidad de enviar señales dirigidas hacia objetivos específicos. En efecto, es esta transmisión específica hacia regiones separadas y muy focalizadas, tanto en el sistema nervioso como en los terminales de los nervios periféricos, lo que le permite llevar a cabo sus incontables funciones de sensibilidad, control motor, memoria y otras muchas.
ANATOMIA FISIOLOGICA DE LA SINAPSIS. una típica motoneurona anterior situada en el asta anterior de la médula espinal. Está compuesta por tres partes fundamentales: el soma, que es el cuerpo principal de la neurona; el único axón, que se extiende desde el soma hacia un nervio periférico para abandonar la médula espinal, y las dendritas, que constituyen una gran cantidad de prolongaciones ramificadas del soma con unas dimensiones hasta de 1 mm de recorrido hacia las zonas adyacentes en la médula.
Sobre la superficie de las dendritas y del soma de la motoneurona se hallan entre 10.000 y 200.000 diminutos botones sinápticos llamados terminales presinápticos, estando aproximadamente del 80 al 95% en las dendritas y solo del 5 al 20% en el soma. Estos terminales presinápticos ocupan el extremo final de las fibrillas nerviosas originadas en muchas otras neuronas. En gran parte son excitadores: es decir, segregan un neurotransmisor que estimula a la neurona postsináptica. Sin embargo, otros son inhibidores, es decir, segregan un neurotransmisor que inhibe a la neurona postsináptica. Las neuronas pertenecientes a otras porciones de la médula y el encéfalo se distinguen de la motoneurona anterior en los siguientes aspectos: 1) las dimensiones del soma celular; 2) la longitud, el tamaño y el número de dendritas, que oscila desde casi cero a muchos centímetros; 3) la longitud y el tamaño del axón, y 4) el número de terminales presinápticos, que puede oscilar desde tan solo unos pocos hasta llegar a 200.000. Estas variaciones hacen que las neuronas situadas en las diversas partes del sistema nervioso reaccionen de forma dispar a las señales sinápticas que les llegan y, por tanto, ejecuten muchas funciones diferentes. Terminales presinápticos Los estudios de los terminales presinápticos efectuados con el microscopio electrónico muestran que poseen variadas formas anatómicas, pero en su mayoría se parecen a pequeños botones redondos u ovalados; de ahí que a veces se les llame botones terminales, botones, pies terminales o botones sinápticos. En él existen dos estructuras internas de importancia para la función excitadora o inhibidora de la sinapsis: las vesículas transmisoras y las mitocondrias. Las vesículas transmisoras contienen el neurotransmisor que, cuando se libera a la hendidura sináptica, excita o inhibe la neurona postsináptica. Excita la neurona postsináptica si la membrana neuronal posee receptores excitadores e inhibe la neurona si contiene receptores inhibidores. Las mitocondrias aportan trifosfato de adenosina (ATP), que a su vez suministra energía para sintetizar más sustancia transmisora. Cuando se propaga un potencial de acción por un terminal presináptico, la despolarización de su membrana hace que una pequeña cantidad de vesículas viertan su contenido hacia la hendidura. Por su parte, el transmisor liberado provoca un cambio inmediato en las características de permeabilidad de la membrana neuronal postsináptica, lo que origina la excitación o la inhibición de la célula, en función de las propiedades del receptor neuronal.
«Mecanismo por el que los potenciales de acción provocan la liberación del transmisor en los terminales presinápticos: misión de los iones calcio« La membrana del terminal presináptico se llama membrana presináptica. Contiene una gran abundancia de canales de calcio dependientes de voltaje. Cuando un potencial de acción la despolariza, estos canales se abren y permiten la entrada en el terminal de un número importante de iones calcio. La cantidad de neurotransmisor que sale a continuación hacia la hendidura sináptica desde el terminal es directamente proporcional al total de iones calcio que penetran. No se conoce el mecanismo exacto por el que estos iones propician su liberación, pero se piensa que es el siguiente: cuando los iones calcio llegan al terminal presináptico, se unen a unas moléculas proteicas especiales situadas sobre la cara interna de la membrana presináptica, llamadas puntos de liberación. A su vez, este enlace suscita la apertura de los puntos de liberación a través de la membrana, y así permite que unas pocas vesículas transmisoras suelten su contenido hacia la hendidura después de cada potencial de acción. En el caso de las vesículas que almacenan el neurotransmisor acetilcolina, existen entre 2.000 y 10.000 moléculas de esta sustancia en cada una y en el terminal presináptico hay suficientes vesículas como para transmitir desde unos cuantos cientos hasta más de 10.000 potenciales de acción. Las moléculas de estos receptores poseen dos elementos importantes: 1) un componente de unión que sobresale fuera desde la membrana hacia la hendidura sináptica y donde se fija el neurotransmisor procedente del terminal presináptico, y 2) un componente intracelular que atraviesa toda la membrana postsináptica hasta el interior de la neurona postsináptica. La activación de los receptores controla la apertura de los canales iónicos en la célula postsináptica según una de las dos formas siguientes: 1) por activación de los canales iónicos para permitir el paso de determinados tipos de iones a través de la membrana, o 2) mediante la activación de un «segundo mensajero» que en vez de un canal iónico es una molécula que protruye hacia el citoplasma celular y activa una sustancia o más en el seno de la neurona postsináptica. A su vez, estos segundos mensajeros aumentan o disminuyen determinadas funciones específicas de la célula. Los receptores de neurotransmisores que activan directamente los canales iónicos a menudo se denominan receptores ionotrópicos, mientras que los que actúan a través de sistemas de segundos mensajeros reciben el nombre de receptores metabotrópicos. Canales iónicos Los canales iónicos de la membrana neuronal postsináptica suelen ser de dos tipos: 1) canales catiónicos, cuya clase más frecuente deja pasar iones sodio cuando se abren, pero a veces también cumplen esta función con el potasio o el calcio, y 2) canales aniónicos, que permiten sobre todo el paso de los iones cloruro, pero también de minúsculas cantidades de otros aniones. Los canales catiónicos que transportan iones sodio están revestidos de cargas negativas. Esta situación atrae hacia ellos a los iones sodio dotados de carga positiva cuando el diámetro del canal aumenta hasta superar el tamaño del ion sodio hidratado. Sin embargo, esas mismas cargas negativas repelen los iones cloruro y otros aniones e impiden su paso. Con respecto a los canales aniónicos, cuando sus diámetros alcanzan las dimensiones suficientes, entran los iones cloruro y los atraviesan hasta el lado opuesto, mientras que los cationes sodio, potasio y calcio quedan retenidos, básicamente porque sus iones hidratados son demasiado grandes para poder pasar.
Sistema de «segundo mensajero» en la neurona postsináptica Muchas funciones del sistema nervioso, como, por ejemplo, los procesos de memoria, requieren la producción de unos cambios prolongados en las neuronas durante segundos y hasta meses después de la desaparición de la sustancia transmisora inicial. Los canales iónicos no son idóneos para originar una variación prolongada en las neuronas postsinápticas, porque se cierran en cuestión de milisegundos una vez desaparece la sustancia transmisora. Sin embargo, en muchos casos, se consigue una excitación o una inhibición neuronal postsináptica a largo plazo al activar un sistema químico de «segundo mensajero» en el interior de esta misma célula, y a continuación será este elemento el que genere el efecto duradero. Existen diversos tipos de sistemas de segundo mensajero. Uno de los más frecuentes recurre a un grupo de proteínas llamadas proteínas G.
Cuando el receptor es activado por un neurotransmisor, después de un impulso nervioso, experimenta un cambio conformacional, que deja expuesto un sitio de unión para el complejo de proteínas G, que después se une a la porción del receptor que sobresale en el interior de la célula. Este proceso permite que la subunidad α libere GDP y, al mismo tiempo, se una al trifosfato de guanosina (GTP) a la vez que separa de las porciones β y γ del complejo. El complejo α-GTP desprendido tiene así libertad de movimiento en el citoplasma celular y ejecuta una función o más entre múltiples posibles, según las características específicas de cada tipo de neurona.
PUEDEN EXISTIE CIERTOS CAMBIOS COMO; Apertura de canales iónicos específicos a través de la membrana celular postsináptica. En el extremo superior derecho de la figura se observa un canal de potasio que está abierto en respuesta a la proteína G; este canal suele permanecer así durante un tiempo prolongado, a diferencia del rápido cierre experimentado por los canales iónicos activados directamente que no recurren al sistema de segundo mensajero. 2. Activación del monofosfato de adenosina cíclico (AMPc) o del monofosfato de guanosina cíclico (GMPc) en la neurona. Recuerde que tanto el AMPc como el GMPc pueden activar una maquinaria metabólica muy específica en la neurona y, por tanto, poner en marcha cualquiera de las múltiples respuestas químicas, entre ellas los cambios prolongados en la propia estructura celular, que a su vez modifican la excitabilidad de la neurona a largo plazo. 3. Activación de una enzima intracelular o más. La proteína G puede activar directamente una enzima intracelular o más. A su vez, las enzimas pueden estimular cualquiera de las numerosas funciones químicas específicas en la célula. 4. Activación de la transcripción génica. La activación de la transcripción génica es uno de los efectos más importantes ocasionados por la activación de los sistemas de segundo mensajero, debido a que la transcripción génica puede provocar la formación de nuevas proteínas en el seno de la neurona, modificando de ese modo su maquinaria metabólica o su estructura. En efecto, se sabe en general que aparecen cambios estructurales en las neuronas oportunamente activadas, sobre todo en los procesos de memoria a largo plazo. La inactivación de la proteína G tiene lugar cuando el GTP unido a la subunidad α se hidroliza para formar GDP. Esta acción lleva a que la subunidad α libere su proteína diana, con lo que inactiva los sistemas de segundos mensajeros, y a continuación vuelve a combinarse con las subunidades β y γ, con lo cual el complejo de proteínas G recupera su estado inactivo. Está claro que la activación de los sistemas de segundo mensajero dentro de la neurona, ya pertenezcan al tipo de la proteína G o a otras clases, resulta importantísima para variar las características de la respuesta a largo plazo en diferentes vías neuronales.
Receptores excitadores o inhibidores en la membrana postsináptica Tras la activación, algunos receptores postsinápticos provocan la excitación de la neurona postsináptica, y otros su inhibición. La importancia de poseer tanto el tipo inhibidor de receptor como el excitador radica en que aporta una dimensión añadida a la función nerviosa, dado que permite tanto limitar su acción como excitarla. Entre los distintos mecanismos moleculares y de membrana empleados por los diversos receptores para provocar la excitación o la inhibición figuran los siguientes. Excitación 1. Apertura de los canales de sodio para dejar pasar grandes cantidades de cargas eléctricas positivas hacia el interior de la célula postsináptica. Esta acción eleva el potencial de membrana intracelular en sentido positivo hasta el nivel umbral para la excitación. Es el medio que se emplea más a menudo con diferencia para ocasionar la excitación. 2. Depresión de la conducción mediante los canales de cloruro, de potasio o ambos. Esta acción reduce la difusión de los iones cloruro con carga negativa hacia el interior de la neurona postsináptica o de los iones potasio con carga positiva hacia el exterior. En cualquier caso, el efecto consiste en volver más positivo de lo normal el potencial de membrana interno, que es excitador. 3. Diversos cambios en el metabolismo interno de la neurona postsináptica para excitar la actividad celular o, en algunas ocasiones, incrementar el número de receptores excitadores de la membrana o disminuir el de los inhibidores. Inhibición 1. Apertura de los canales del ion cloruro en la membrana neuronal postsináptica. Esta acción permite la difusión rápida de iones cloruro dotados de carga negativa desde el exterior de la neurona postsináptica hacia su interior, lo que traslada estas cargas al interior y aumenta la negatividad en esta zona, efecto que tiene un carácter inhibidor. 2. Aumento de la conductancia para los iones potasio fuera de la neurona. Esta acción permite la difusión de iones positivos hacia el exterior, lo que causa una mayor negatividad dentro de la neurona; esto representa una acción inhibidora. 3. Activación de las enzimas receptoras que inhiben las funciones metabólicas celulares encargadas de aumentar el número de receptores sinápticos inhibidores o de disminuir el de los excitadores.
«TRANSMISORES DE ACCIÓN RÁPIDA Y MÓLECULA PEQUEÑA.» En la mayoría de los casos, los tipos de transmisores de molécula pequeña se sintetizan en el citoplasma del terminal presináptico y las numerosas vesículas transmisoras presentes a este nivel los absorben por transporte activo. Reciclado de las vesículas de molécula pequeña Las vesículas que se almacenan y liberan transmisores de molécula pequeña se reciclan continuamente y se utilizan una y otra vez. Una vez que se fusionan con la membrana sináptica y se abren para verter la sustancia transmisora, la membrana de la vesícula simplemente forma parte al principio de la membrana sináptica.
La acetilcolina es un típico transmisor de molécula pequeña que obedece a los principios de síntesis y liberación antes expuestos. Esta sustancia transmisora se sintetiza en el terminal presináptico a partir de acetil coenzima A y colina en presencia de la enzima acetiltransferasa de colina. A continuación, se transporta a sus vesículas específicas. Cuando más tarde se produce su salida desde ellas a la hendidura sináptica durante la transmisión de la señal nerviosa en la sinapsis, se degrada de nuevo con rapidez en acetato y colina por acción de la enzima colinesterasa, que está presente en el retículo formado por proteoglucano que rellena el espacio de la hendidura sináptica. Después, una vez más, las vesículas se reciclan en el interior del terminal presináptico, y la colina sufre un transporte activo de vuelta hacia el terminal para repetir su empleo en la síntesis de nueva acetilcolina. Características de algunos importantes transmisores de molécula pequeña La acetilcolina se segrega por las neuronas situadas en muchas regiones del sistema nervioso, pero específicamente en: 1) los terminales de las células piramidales grandes de la corteza motora; 2) diversos tipos diferentes de neuronas pertenecientes a los ganglios basales; 3) las motoneuronas que inervan los músculos esqueléticos; 4) las neuronas preganglionares del sistema nervioso autónomo; 5) las neuronas posganglionares del sistema nervioso parasimpático, y 6) parte de las neuronas posganglionares del sistema nervioso simpático. En la mayoría de los casos, la acetilcolina posee un efecto excitador; sin embargo, se sabe que ejerce acciones inhibidoras en algunas terminaciones nerviosas parasimpáticas periféricas, como la inhibición del corazón a cargo de los nervios vagos. La noradrenalina se segrega en los terminales de muchas neuronas cuyos somas están situados en el tronco del encéfalo y el hipotálamo. En concreto, las que están localizadas en el locus ceruleus de la protuberancia envían fibras nerviosas a amplias regiones del encéfalo que sirven para controlar la actividad global y el estado mental, como por ejemplo aumentar el nivel de vigilia. En la mayoría de estas zonas, la noradrenalina probablemente activa receptores excitadores, pero en unas cuantas, en cambio, estimula los inhibidores. También se segrega en la mayor parte de las neuronas posganglionares del sistema nervioso simpático, donde excita algunos órganos pero inhibe otros. La dopamina se segrega en las neuronas originadas en la sustancia negra. Su terminación se produce básicamente en la región estriada de los ganglios basales. El efecto que ejerce suele ser una inhibición. La glicina se segrega sobre todo en las sinapsis de la médula espinal. Se cree que siempre actúa como un transmisor inhibidor. El GABA (ácido γ-aminobutírico) se segrega en los terminales nerviosos de la médula espinal, el cerebelo, los ganglios basales y muchas áreas de la corteza. Se piensa que siempre causa una inhibición. El glutamato se segrega en los terminales presinápticos de muchas de las vías sensitivas que penetran en el sistema nervioso central, lo mismo que en muchas áreas de la corteza cerebral. Probablemente siempre causa excitación. La serotonina se segrega en los núcleos originados en el rafe medio del tronco del encéfalo que proyectan hacia numerosas regiones del cerebro y de la médula espinal, especialmente a las astas dorsales de la médula y al hipotálamo. Actúa en la médula como un inhibidor de las vías del dolor, y se piensa que la acción inhibidora sobre las regiones superiores del sistema nervioso ayuda a controlar el estado de ánimo de una persona, tal vez incluso provocando sueño. El óxido nítrico se segrega especialmente en los terminales nerviosos de las regiones encefálicas responsables de la conducta a largo plazo y de la memoria. Por tanto, este sistema transmisor podría esclarecer en el futuro algunas de las funciones correspondientes a estos dos aspectos que hasta ahora han desafiado toda explicación. El óxido nítrico difiere de otros transmisores de molécula pequeña por su mecanismo de producción en el terminal presináptico y por sus acciones sobre la neurona postsináptica. No está formado con antelación y almacenado en vesículas dentro del terminal presináptico como los demás transmisores. En su lugar, se sintetiza casi al instante según las necesidades, y a continuación difunde fuera de los terminales presinápticos durante un período de segundos en vez de ser liberado en paquetes vesiculares, y después hacia las neuronas postsinápticas cercanas. En ellas, no suele alterar mucho el potencial de membrana, sino que modifica las funciones metabólicas intracelulares que cambian la excitabilidad neuronal durante segundos, minutos o tal vez incluso más tiempo. Neuropéptidos Los neuropéptidos se sintetizan de otro modo y tienen acciones que normalmente son lentas y en otros aspectos bastante diferentes de las que ejercen los transmisores de molécula pequeña. Estas sustancias no se sintetizan en el citoplasma de los terminales presinápticos. Por el contrario, se forman en los ribosomas del soma neuronal ya como porciones íntegras de grandes moléculas proteicas. Las moléculas proteicas penetran a continuación en los espacios existentes en el retículo endoplásmico del soma y posteriormente en el aparato de Golgi, donde suceden dos cambios. En primer lugar, la proteína formadora de neuropéptidos sufre una escisión enzimática en fragmentos más pequeños, algunos de los cuales son el propio neuropéptido o un precursor suyo. En segundo lugar, el aparato de Golgi introduce el neuropéptido en minúsculas vesículas transmisoras que se liberan hacia el citoplasma. A continuación, se transportan por el axón en todas las direcciones hacia el extremo de las fibras nerviosas a través de la corriente axónica del citoplasma, viajando a una velocidad de tan solo unos pocos centímetros al día. Finalmente, estas vesículas vierten su contenido en los terminales neuronales como respuesta a los potenciales de acción de la misma manera que los transmisores de molécula pequeña. Sin embargo, la vesícula sufre una autólisis y no se reutiliza. Debido a este laborioso método de formación de los neuropéptidos, normalmente se libera una cantidad mucho menor de neuropéptidos que de los transmisores de molécula pequeña. Esta diferencia se compensa en parte por el hecho de que en general poseen una potencia 1.000 veces mayor o más que los transmisores de molécula pequeña. Otra característica importante de los neuropéptidos es que a menudo ocasionan acciones mucho más duraderas. Algunas de ellas consisten en el cierre prolongado de los canales de calcio, los cambios persistentes en la maquinaria metabólica de las células, en la activación o la desactivación de genes específicos dentro del núcleo celular, o en alteraciones a largo plazo de la cantidad de receptores excitadores o inhibidores. Algunos de estos efectos duran días, pero otros quizá meses o años. Nuestros conocimientos sobre las funciones de los neuropéptidos no están sino comenzando a desarrollarse. Fenómenos eléctricos durante la excitación neuronal Los fenómenos eléctricos que ocurren durante la excitación neuronal se han estudiado especialmente en las grandes motoneuronas situadas en las astas anteriores de la médula espinal. Por tanto, los sucesos descritos en los próximos apartados se refieren básicamente a estas neuronas. Excepto por sus diferencias cuantitativas, también pueden aplicarse a la mayoría de las demás neuronas del sistema nervioso.
POTENCIAL DE MEMBRANA EN REPOSO DEL SOMA NEURONAL.
Este potencial de membrana en reposo es un poco menos negativo que los – 90 mV existentes en las grandes fibras nerviosas periféricas y en las del músculo esquelético; un voltaje más bajo resulta importante ya que permite el control positivo y negativo del grado de excitabilidad neuronal. Es decir, el descenso del voltaje hasta un nivel menos negativo vuelve más excitable la membrana de la neurona, mientras que su aumento hasta un nivel más negativo la hace menos excitable. Este mecanismo es el fundamento de los dos modos de acción en la neurona, la excitación o la inhibición.
El interior del soma neuronal contiene una solución electrolítica muy conductora, el líquido intracelular de la neurona. Además, su diámetro es grande (de 10 a 80 μm), lo que casi no genera ninguna resistencia a la conducción de la corriente eléctrica de una parte a otra de su interior. Por tanto, todo cambio en el potencial de cualquier zona del líquido dentro del soma suscita un cambio casi exactamente igual en el potencial de los demás puntos de su interior (esto es, siempre que la neurona no esté transmitiendo un potencial de acción). Este principio es importante, porque desempeña un cometido fundamental en la «sumación» de las señales que llegan a la neurona desde múltiples fuentes
Generación de potenciales de acción en el segmento inicial del axón a su salida de la neurona: umbral de excitación Cuando el PPSE sube lo suficiente en sentido positivo, llega a un punto en el que pone en marcha un potencial de acción en la neurona. Sin embargo, este potencial no empieza en las inmediaciones de las sinapsis excitadoras. En su lugar, empieza en el segmento inicial del axón al nivel en que esta estructura abandona el soma neuronal. La razón principal para que este sea el punto de origen del potencial de acción reside en que el soma posee en su membrana relativamente pocos canales de sodio dependientes de voltaje, lo que complica la apertura por parte del PPSE del número necesario para desencadenar un potencial de acción. Por el contrario, la membrana del segmento inicial presenta una concentración siete veces superior que el soma de canales de sodio dependientes de voltaje y, por tanto, puede generar un potencial de acción con mucha mayor facilidad que este último. El PPSE que suscitará un potencial de acción en el segmento inicial del axón está entre +10 y +20 mV, en contraste con los +30 o +40 mV requeridos como mínimo en el soma. POTENCIAL DE MEMBRANA EN REPOSO DEL SOMA NEURONAL.
Este potencial de membrana en reposo es un poco menos negativo que los – 90 mV existentes en las grandes fibras nerviosas periféricas y en las del músculo esquelético; un voltaje más bajo resulta importante ya que permite el control positivo y negativo del grado de excitabilidad neuronal. Es decir, el descenso del voltaje hasta un nivel menos negativo vuelve más excitable la membrana de la neurona, mientras que su aumento hasta un nivel más negativo la hace menos excitable. Este mecanismo es el fundamento de los dos modos de acción en la neurona, la excitación o la inhibición.
El interior del soma neuronal contiene una solución electrolítica muy conductora, el líquido intracelular de la neurona. Además, su diámetro es grande (de 10 a 80 μm), lo que casi no genera ninguna resistencia a la conducción de la corriente eléctrica de una parte a otra de su interior. Por tanto, todo cambio en el potencial de cualquier zona del líquido dentro del soma suscita un cambio casi exactamente igual en el potencial de los demás puntos de su interior (esto es, siempre que la neurona no esté transmitiendo un potencial de acción). Este principio es importante, porque desempeña un cometido fundamental en la «sumación» de las señales que llegan a la neurona desde múltiples fuentes.
Fenómenos eléctricos durante la inhibición neuronal Efecto de las sinapsis inhibidoras sobre la membrana postsináptica: potencial postsináptico inhibidor Las sinapsis inhibidoras sobre todo abren canales de cloruro, lo que permite el paso sin problemas de estos iones. Para comprender cómo inhiben la neurona postsináptica, debemos recordar lo que aprendimos sobre el potencial de Nernst para los iones cloruro. Calculamos que en este caso dicha variable mide unos –70 mV. Se trata de un potencial más negativo que los –65 mV presentes normalmente en el interior de la membrana neuronal en reposo. Por tanto, la apertura de los canales de cloruro permitirá el movimiento de estos iones con carga negativa desde el líquido extracelular hacia el interior, lo que volverá más negativo de lo normal el potencial de membrana interno, acercándolo al nivel de –70 mV.
Inhibición presináptica Además de la inhibición originada por las sinapsis inhibidoras que operan en la membrana neuronal, lo que se denomina inhibición postsináptica, muchas veces se produce otro tipo de inhibición en los terminales presinápticos antes de que la señal llegue a alcanzar la sinapsis. Esta clase se llama inhibición presináptica. La inhibición presináptica está ocasionada por la liberación de una sustancia inhibidora en las inmediaciones de las fibrillas nerviosas presinápticas antes de que sus propias terminaciones acaben sobre la neurona postsináptica. En la mayoría de los casos, la sustancia transmisora inhibidora es GABA (ácido γ-aminobutírico). Esta liberación ejerce una acción específica de apertura sobre los canales aniónicos, lo que permite la difusión de una gran cantidad de iones cloruro hacia la fibrilla terminal. Las cargas negativas de estos iones inhiben la transmisión sináptica debido a que anulan gran parte del efecto excitador producido por los iones sodio con carga positiva que también penetran en las fibrillas terminales cuando llega un potencial de acción. La inhibición presináptica ocurre en muchas de las vías sensitivas del sistema nervioso. En realidad, las fibras nerviosas sensitivas adyacentes entre sí suelen inhibirse mutuamente, lo que atenúa la propagación lateral y la mezcla de señales en los fascículos sensitivos.
«Sumación espacial» en las neuronas: umbral de disparo La excitación de un solo terminal presináptico sobre la superficie de una neurona casi nunca activa la célula. La razón de este fenómeno reside en que la cantidad de sustancia transmisora liberada por un terminal aislado para originar un PPSE normalmente no supera los 0,5 a 1 mV, en vez de los 10 a 20 mV necesarios en general para alcanzar el umbral de excitación. Sin embargo, al mismo tiempo suelen estimularse muchos terminales presinápticos. Aunque estas estructuras se encuentren esparcidas por amplias regiones de la neurona, aun así pueden sumarse sus efectos; es decir, agregarse uno a otro hasta que se produzca la excitación neuronal. Ya se señaló antes que un cambio de potencial en cualquier punto aislado del soma provocará su modificación casi exactamente igual hacia todas partes en su interior. Este efecto es así debido a que el gran cuerpo de la neurona presenta una conductividad eléctrica muy alta. Por tanto, con cada sinapsis excitadora que descarga a la vez, el potencial total dentro del soma se vuelve más positivo en 0,5 a 1 mV. Cuando el PPSE llegue al nivel suficiente, alcanzará el umbral de disparo y producirá un potencial de acción espontáneamente en el segmento inicial del axón
Cada vez que dispara un terminal presináptico, la sustancia transmisora liberada abre los canales de membrana a lo sumo durante 1 ms más o menos. Sin embargo, la modificación del potencial postsináptico dura hasta 15 ms una vez que los canales de membrana sinápticos ya están cerrados. Por tanto, una segunda apertura de estos mismos elementos puede incrementar el potencial postsináptico hasta un nivel aún mayor y cuanto más alta sea la velocidad de estimulación, mayor se volverá el potencial postsináptico. Así pues, las descargas sucesivas de un solo terminal presináptico, si suceden con la rapidez suficiente, pueden añadirse unas a otras; es decir, pueden «sumarse». Este tipo de adición se denomina sumación temporal.
«Facilitación» de las neuronas Con frecuencia el potencial postsináptico total una vez sumado es excitador, pero no ha subido lo suficiente como para alcanzar el umbral de disparo en la neurona postsináptica. Cuando se produce esta situación, se dice que la neurona está facilitada. Es decir, su potencial de membrana está más cerca del umbral de disparo que lo normal, pero aún no ha alcanzado este nivel. Por consiguiente, la llegada de una señal excitadora más emanada de cualquier otra fuente puede activarla en estas condiciones con una gran facilidad. Las señales difusas del sistema nervioso suelen facilitar grandes grupos de neuronas para que sean capaces de responder con rapidez y sin problemas a las señales que dimanan de otros orígenes.
Funciones especiales de las dendritas para excitar a las neuronas Campo espacial de excitación de las dendritas amplio Las dendritas de las motoneuronas anteriores suelen extenderse de 500 a 1.000 μm en todas las direcciones a partir del soma neuronal, y pueden recibir señales procedentes de una gran región espacial en torno a la motoneurona. Esta característica ofrece enormes oportunidades para su sumación desde numerosas fibras nerviosas presinápticas independientes. También es importante resaltar que entre el 80 y el 95% de todos los terminales presinápticos de la motoneurona anterior acaban sobre dendritas, a diferencia de un mero 5-20% que finalizan sobre el soma neuronal. Por consiguiente, una gran parte de la excitación viene suministrada por las señales transmitidas a través de las dendritas. La mayoría de las dendritas no son capaces de transmitir potenciales de acción, pero sí señales dentro de la misma neurona mediante conducción electrotónica La mayoría de las dendritas no llegan a transmitir potenciales de acción debido a que sus membranas poseen relativamente pocos canales de sodio dependientes de voltaje, y sus umbrales de excitación son demasiado elevados para producir potenciales de acción. Con todo, sí que transportan corrientes electrotónicas desde las dendritas al soma. Este proceso significa la propagación directa de una corriente eléctrica por conducción de iones en los líquidos de las dendritas pero sin la generación de potenciales de acción. La estimulación (o inhibición) de la neurona por esta corriente presenta unas características especiales
En las dos dendritas de la izquierda aparecen efectos excitadores cerca del extremo de su punta; obsérvese el elevado nivel de los PPSE en estas porciones, es decir, fíjese en sus potenciales de membrana menos negativos en estos puntos. Sin embargo, una gran parte de su PPSE se pierde antes de llegar al soma. La razón de esta importante pérdida estriba en que las dendritas son largas y sus membranas delgadas y al menos parcialmente permeables a los iones potasio y cloruro, lo que las vuelve «porosas» a la corriente eléctrica. Por tanto, antes de que los potenciales excitadores puedan alcanzar el soma, una gran proporción se habrá perdido por escape a través de la membrana. Esta disminución del potencial de membrana a medida que experimenta su propagación electrotónica a lo largo de las dendritas hacia el soma se llama conducción decreciente.
Algunas neuronas del sistema nervioso central disparan de forma continua porque incluso su estado excitador normal se encuentra por encima del nivel umbral. La frecuencia de disparo suele poder elevarse aún más con un nuevo incremento de su estado excitador. En cambio, puede reducirse, o incluso detenerse los disparos, si se superpone un estado inhibidor en la neurona. Por tanto, dos neuronas distintas responden de modos diferentes, tienen umbrales de excitación dispares y presentan
unas frecuencias máximas de descarga muy alejadas entre sí. Con un poco de imaginación es posible comprender fácilmente la importancia de poseer neuronas diversas con unas características de respuesta de múltiples tipos para cumplir las funciones muy variadas del sistema nervioso.
Algunas características especiales de la transmisión sináptica Fatiga de la transmisión sináptica Cuando las sinapsis excitadoras reciben estímulos repetidos a un ritmo elevado, el número de descargas de la neurona postsináptica es muy alto al principio, pero la frecuencia de disparo va bajando progresivamente en los milisegundos o segundos sucesivos. Este fenómeno se llama fatiga de la transmisión sináptica. La fatiga es una característica importantísima de la función sináptica porque cuando una región del sistema nervioso está hiperexcitada, permite que desaparezca este exceso de excitabilidad pasado un rato. Por ejemplo, la fatiga probablemente es el medio más sobresaliente para acabar dominando la excitabilidad excesiva del encéfalo durante una crisis epiléptica, hasta el punto de que cesen las convulsiones. Por tanto, su aparición constituye un mecanismo protector contra el exceso de actividad neuronal.
Efecto de la acidosis o de la alcalosis sobre la transmisión sináptica La mayoría de las neuronas son muy sensibles a los cambios del pH en los líquidos intersticiales que las rodean. Normalmente, la alcalosis aumenta mucho la excitabilidad neuronal. Por ejemplo, un ascenso en el pH de la sangre arterial desde su valor habitual de 7,4 hasta 7,8 u 8 suele causar convulsiones epilépticas en el encéfalo debido a la mayor excitabilidad de algunas neuronas cerebrales o de todas. En una persona predispuesta a convulsiones epilépticas, incluso un período breve de hiperventilación, que elimina el dióxido de carbono y eleva el pH de la sangre, puede desencadenar un ataque epiléptico. En cambio, la acidosis disminuye acusadamente la actividad neuronal; un descenso en el pH desde 7,4 hasta un valor inferior a 7 suele ocasionar un estado comatoso. Por ejemplo, en la acidosis diabética o urémica muy grave, casi siempre se presenta coma.
Efecto de la hipoxia sobre la transmisión sináptica La excitabilidad neuronal también depende claramente de un aporte suficiente de oxígeno. Su interrupción por unos pocos segundos puede ocasionar una ausencia completa de excitabilidad en algunas neuronas. Este efecto se observa cuando cesa transitoriamente el flujo sanguíneo cerebral, porque en cuestión de 3 a 7 s la persona pierde el conocimiento. Efecto de los fármacos sobre la transmisión sináptica Se sabe que muchos fármacos aumentan la excitabilidad de las neuronas y otros la disminuyen. Por ejemplo, la cafeína, la teofilina y la teobromina, que están presentes en el café, el té y el chocolate, respectivamente, incrementan la excitabilidad neuronal, se supone que al rebajar el umbral de excitación en las células. La estricnina es uno de los productos mejor conocidos que aumenta la excitabilidad de las neuronas. Sin embargo, no lo hace reduciendo su umbral de excitación; en su lugar, inhibe la acción de algunas sustancias transmisoras normalmente inhibidoras, sobre todo el efecto en este sentido de la glicina sobre la médula espinal. Por tanto, las acciones de los transmisores excitadores resultan aplastantes, y las neuronas quedan tan excitadas que pasan con rapidez a emitir descargas repetidas, derivando en unos espasmos musculares tónicos de gran intensidad. La mayoría de los anestésicos elevan el umbral de la membrana neuronal para la excitación y así disminuyen la transmisión sináptica en muchos puntos del sistema nervioso. Como muchos de estos compuestos son especialmente liposolubles, se ha pensado que algunos de ellos podrían modificar las propiedades físicas de las membranas neuronales, volviéndolas menos sensibles a los productos excitadores. Retraso sináptico Durante la transmisión de una señal neuronal desde una neurona presináptica hasta otra postsináptica, se consume cierta cantidad de tiempo en el proceso siguiente: 1) emisión de la sustancia transmisora por el terminal presináptico; 2) difusión del transmisor hacia la membrana neuronal postsináptica; 3) acción del transmisor sobre el receptor de la membrana; 4) intervención del receptor para aumentar la permeabilidad de la membrana, y 5) entrada del sodio por difusión para elevar el PPSE hasta un nivel suficientemente alto como para desencadenar un potencial de acción. El período mínimo necesario para que tengan lugar todos estos fenómenos, incluso cuando se estimula simultáneamente un gran número de sinapsis excitadoras, es de unos 0,5 ms, que se denomina retraso sináptico. Los neurofisiólogos pueden medir el tiempo de retraso mínimo transcurrido entre la llegada de una lluvia de impulsos a un conjunto de neuronas y la correspondiente lluvia de salida. Una vez recogido este dato, ya se puede calcular el número de neuronas sucesivas que forman el circuito.
El músculo liso, que está formado por fibras mucho menores, habitualmente de 1 a 5 μm de diámetro y de solo 20 a 500 μm de longitud. <«TIPOS DE MUSCULO LISO»> El músculo liso de los distintos órganos es distinto del de la mayor parte de los demás en varios sentidos: 1) dimensiones físicas; 2) organización en fascículos o láminas; 3) respuesta a diferentes tipos de estímulos; 4) características de la inervación, y 5) función. Músculo liso multiunitario. Este tipo de músculo liso está formado por fibras musculares lisas separadas y discretas. Cada una de las fibras actúa independientemente de las demás y con frecuencia está inervada por una única terminación nerviosa, como ocurre en las fibras musculares esqueléticas. Además, la superficie externa de estas fibras, como en el caso de las fibras musculares esqueléticas, está cubierta por una capa delgada de sustancia similar a una membrana basal, una mezcla de colágeno fino y glucoproteínas que aísla las fibras separadas entre sí. Las características fundamentales de las fibras musculares lisas multiunitarias son que cada una de las fibras se puede contraer independientemente de las demás, y que su control se ejerce principalmente por señales nerviosas. Algunos ejemplos de músculo liso multiunitario son el músculo ciliar del ojo, el músculo del iris del ojo y los músculos piloerectores que producen la erección del pelo cuando los estimula el sistema nervioso simpático.
<«MUSCULO LISO UNITARIO»> Este tipo se denomina músculo liso sincitial o músculo liso visceral. El término «unitario» es confuso porque no se refiere a fibras musculares únicas. Las fibras habitualmente están dispuestas en láminas o fascículos, y sus membranas celulares están adheridas entre sí en múltiples puntos, de modo que la fuerza que se genera en una fibra muscular se puede transmitir a la siguiente. las membranas celulares están unidas por muchas uniones en hendidura a través de las cuales los iones pueden fluir libremente desde una célula muscular a otra, de modo que los potenciales de acción o el flujo iónico simple sin potenciales de acción puede viajar desde una fibra a otra y hacer que las fibras musculares se contraigan simultáneamente. Este tipo de músculo liso también se conoce como músculo liso sincitial debido a sus interconexiones sincitiales entre las fibras. También se denomina músculo liso visceral porque se encuentra en la pared de la mayor parte de las vísceras del cuerpo, por ejemplo el aparato digestivo, las vías biliares, los uréteres, el útero y muchos vasos sanguíneos.
Base química de la contracción del músculo liso El músculo liso contiene filamentos tanto de actina como de miosina, que tienen características químicas similares a los filamentos de actina y miosina del músculo esquelético. No contiene el complejo de troponina necesario para el control de la contracción del músculo esquelético, de modo que el mecanismo de control de la contracción es diferente. Estudios químicos han mostrado que los filamentos de actina y miosina del músculo liso interactúan entre sí de manera muy similar a como lo hacen en el músculo esquelético. Además, el proceso contráctil es activado por los iones calcio, y el trifosfato de adenosina (ATP) se degrada a difosfato de adenosina (ADP) para proporcionar la energía para la contracción. El músculo liso no tiene la misma disposición estriada de los filamentos de actina y miosina que se encuentra en el músculo esquelético Interpuestos entre los filamentos de actina de la fibra muscular están los filamentos de miosina. Estos filamentos tienen un diámetro superior al doble que los filamentos de actina. En las microfotografías electrónicas habitualmente se ven 5 a 10 veces más filamentos de actina que de miosina, la mayoría de los filamentos de miosina tienen lo que se denomina puentes cruzados «lateropolares», dispuestos de tal manera que los puentes de un lado basculan en una dirección y los del otro lado basculan en la dirección opuesta. Esta configuración permite que la miosina tire de un filamento de actina en una dirección en un lado a la vez que tira de otro filamento de actina en la dirección opuesta en el otro lado, la mayoría de los músculos esqueléticos se contraen y relajan rápidamente, muchas de las contracciones del músculo liso son contracciones tónicas prolongadas, que a veces duran horas o incluso días. Por tanto, cabe esperar que las características físicas y químicas de la contracción del músculo liso sean diferentes de las del músculo esquelético.
PUENTES CRUZADOS DE LA MIOSINA. La rapidez del ciclado de los puentes transversos de miosina en el músculo liso (es decir, su unión a la actina, su posterior liberación de la actina y su nueva unión para el siguiente ciclo) es mucho más lenta que en el músculo esquelético; de hecho, la frecuencia es tan baja como 1/10 a 1/300 de la del músculo esquelético, Una posible razón del ciclado lento es que las cabezas de los puentes cruzados tienen una actividad ATPasa mucho menor que en el músculo esquelético, de modo que la degradación del ATP que aporta energía a los movimientos de las cabezas de los puentes cruzados está muy reducida, con el consiguiente enlentecimiento de la frecuencia de ciclado. También se piensa que esto se debe al lento ciclado de unión y separación de los puentes cruzados y a que solo es necesaria una molécula de ATP para cada ciclo, independientemente de su duración. La baja utilización de energía por el músculo liso es importante para la economía energética global del cuerpo, porque órganos como los intestinos, la vejiga urinaria, la vesícula biliar y otras vísceras con frecuencia mantienen una contracción muscular tónica casi indefinidamente.
El inicio lento de la contracción del músculo liso, así como su contracción prolongada, está producido por la lentitud de la unión y la separación de los puentes cruzados a los filamentos de actina. Además, el inicio de la contracción en respuesta a los iones calcio es mucho más lento que en el músculo esquelético, a pesar de la escasez relativa de filamentos de miosina en el músculo liso, y a pesar del tiempo lento de ciclado de los puentes cruzados, la fuerza máxima de contracción del músculo liso es con frecuencia mayor que la del músculo esquelético, hasta 4 a 6 kg/cm2 de área transversal para el músculo liso, en comparación con 3 a 4 kg para el músculo esquelético. TENSIÓN-RELAJACIÓN DEL MÚSCULO LISO. Otra característica importante del músculo liso, especialmente del tipo unitario visceral de músculo liso de muchos órganos huecos, es su capacidad de recuperar casi su fuerza de contracción original segundos a minutos después de que haya sido alargado o acortado, cuando se produce una reducción súbita de volumen, la presión disminuye drásticamente al principio, aunque después aumenta en un plazo de otros pocos segundos o minutos hasta o casi hasta el nivel original. Estos fenómenos se denominan tensión-relajación y tensiónrelajación inversa. Su importancia radica en que, excepto durante breves períodos, permiten que un órgano hueco mantenga aproximadamente la misma presión en el interior de su luz a pesar de grandes cambios de volumen sostenidos.
Regulación de la contracción por los iones calcio.
El músculo liso no contiene troponina, la proteína reguladora que es activada por los iones calcio para producir la contracción del músculo esquelético. En cambio, la contracción del músculo liso está activada por un mecanismo totalmente distinto, como se describe en el siguiente apartado, en lugar de la troponina, las células musculares lisas contienen una gran cantidad de otra proteína reguladora denominada calmodulina . Esta activación y la posterior contracción se producen según la siguiente secuencia: 1. La concentración de calcio en el líquido citosólico del músculo liso se incrementa como consecuencia de la entrada de calcio desde el líquido extracelular a través de los canales de calcio y/o la liberación de calcio desde el retículo sarcoplásmico. 2. Los iones calcio se unen a la calmodulina de forma reversible. 3. El complejo calmodulina-calcio se une después a la miosina cinasa de cadena ligera, que es una enzima fosforiladora, y la activa. 4. Una de las cadenas ligeras de cada una de las cabezas de miosina, denominada cabeza reguladora, se fosforila en respuesta a esta miosina cinasa. Cuando esta cadena no está fosforilada no se produce el ciclo de unión-separación de la cabeza de miosina con el filamento de actina. Sin embargo, cuando la cadena reguladora está fosforilada la cabeza tiene la capacidad de unirse repetitivamente al filamento de actina y de avanzar a través de todo el proceso de ciclado de «tirones» intermitentes, al igual que ocurre en el músculo esquelético, produciendo de esta manera la contracción muscular.
Aunque el proceso contráctil en el músculo liso, como en el músculo esquelético, es activado por los iones calcio, el origen de estos iones es diferente. Una distinción importante es que el retículo sarcoplásmico, que proporciona prácticamente todos los iones calcio para la contracción musculoesquelética, está desarrollado solo ligeramente en la mayor parte del músculo liso. la concentración de iones calcio en el líquido extracelular es superior a 10−3 molar, en comparación con un valor inferior a 10−7 molar en el interior de la célula muscular lisa; esta situación origina una rápida difusión de los iones calcio en la célula desde el líquido extracelular cuando se abren los canales de calcio. El tiempo necesario para que tenga lugar esta difusión se sitúa, en promedio, entre 200 y 300 ms, y recibe el nombre de período latente, antes de que se inicie la contracción. Aunque el proceso contráctil en el músculo liso, como en el músculo esquelético, es activado por los iones calcio, el origen de estos iones es diferente. Una distinción importante es que el retículo sarcoplásmico, que proporciona prácticamente todos los iones calcio para la contracción musculoesquelética, está desarrollado solo ligeramente en la mayor parte del músculo liso, algunos retículos sarcoplásmicos muy poco desarrollados que se sitúan cerca de las membranas celulares en algunas células del músculo liso más grandes. Pequeñas invaginaciones de la membrana celular, denominadas cavéolas, terminan en las superficies de estos túbulos. Las cavéolas sugieren una rudimentaria analogía del sistema de túbulos transversos del músculo esquelético. Cuando se transmite un potencial de acción a las cavéolas, según se cree estimula la liberación de iones calcio desde los túbulos sarcoplásmicos contiguos de la misma forma que los potenciales de acción en los túbulos transversos del músculo esquelético provocan la liberación de iones calcio desde los túbulos sarcoplásmicos transversosAunque el proceso contráctil en el músculo liso, como en el músculo esquelético, es activado por los iones calcio, el origen de estos iones es diferente. Una distinción importante es que el retículo sarcoplásmico, que proporciona prácticamente todos los iones calcio para la contracción musculoesquelética, está desarrollado solo ligeramente en la mayor parte del músculo liso. la concentración de iones calcio en el líquido extracelular es superior a 10−3 molar, en comparación con un valor inferior a 10−7 molar en el interior de la célula muscular lisa; esta situación origina una rápida difusión de los iones calcio en la célula desde el líquido extracelular cuando se abren los canales de calcio. El tiempo necesario para que tenga lugar esta difusión se sitúa, en promedio, entre 200 y 300 ms, y recibe el nombre de período latente, antes de que se inicie la contracción. Aunque el proceso contráctil en el músculo liso, como en el músculo esquelético, es activado por los iones calcio, el origen de estos iones es diferente. Una distinción importante es que el retículo sarcoplásmico, que proporciona prácticamente todos los iones calcio para la contracción musculoesquelética, está desarrollado solo ligeramente en la mayor parte del músculo liso.
La miosina fosfatasa es importante en la interrupción de la contracción La relajación del músculo liso tiene lugar cuando los canales de calcio se cierran y la bomba de calcio transporta iones calcio fuera del líquido citosólico de la célula. Cuando la concentración de iones calcio disminuye por debajo de un nivel crítico, los procesos que se acaban de señalar se invierten automáticamente, excepto la fosforilación de la cabeza de miosina. Cuando las enzimas miosina cinasa y miosina fosfatasa están intensamente activadas, la frecuencia de ciclado de las cabezas de miosina y la velocidad de contracción son elevadas. Después, cuando disminuye la activación de las enzimas, lo hace también la frecuencia de ciclado, pero al mismo tiempo la desactivación de estas enzimas permite que las cabezas de miosina permanezcan unidas al filamento de actina durante una proporción cada vez mayor del período de ciclado. Control nervioso y hormonal de la contracción del músculo liso Aunque las fibras musculares esqueléticas son estimuladas exclusivamente por el sistema nervioso, la contracción del músculo liso puede ser estimulada por señales nerviosas, estimulación hormonal, distensión del músculo y otros diversos estímulos. El principal motivo de esta diferencia es que la membrana del músculo liso contiene muchos tipos de proteínas receptoras que pueden iniciar el proceso contráctil. Además, otras proteínas receptoras inhiben la contracción del músculo liso, que es otra diferencia respecto al músculo esquelético. Las uniones neuromusculares del tipo muy estructurado que se encuentran en las fibras del músculo esquelético no aparecen en el músculo liso, las fibras nerviosas autónomas que inervan el músculo liso generalmente se ramifican de manera difusa encima de una lámina de fibras musculares. Los axones que inervan las fibras musculares lisas no tienen los extremos terminales ramificados típicos que se observan en la placa motora terminal de las fibras musculares esqueléticas. Por el contrario, la mayoría de los axones terminales delgados tiene múltiples varicosidades distribuidas a lo largo de sus ejes. En estos puntos se interrumpen las células de Schwann que rodean a los axones, de modo que se puede secretar la sustancia transmisora a través de las paredes de las varicosidades. En las varicosidades hay vesículas similares a las de la placa terminal del músculo esquelético y que contienen la sustancia transmisora. Las sustancias transmisoras más importantes que secretan los nervios autónomos que inervan el músculo liso son acetilcolina y noradrenalina, aunque nunca son secretadas por las mismas fibras nerviosas. La acetilcolina es una sustancia transmisora excitadora de las fibras musculares lisas en algunos órganos y un transmisor inhibidor en el músculo liso de otros órganos. Cuando la acetilcolina excita una fibra, la noradrenalina habitualmente la inhibe.
Potenciales de membrana en el músculo liso El voltaje cuantitativo del potencial de membrana del músculo liso depende de la situación momentánea del músculo. En el estado de reposo normal el potencial intracelular es de aproximadamente –50 a –60 mV, alrededor de 30 mV menos negativo que en el músculo esquelético. Potenciales de acción en el músculo liso unitario Los potenciales de acción se producen en el músculo liso unitario (como el músculo visceral) de la misma forma que en el músculo esquelético.
La membrana de la célula muscular lisa tiene muchos más canales de calcio activados por el voltaje que el músculo esquelético, aunque tiene pocos canales de sodio activados por el voltaje. Por tanto, el sodio participa poco en la generación del potencial de acción en la mayor parte del músculo liso. Por el contrario, el flujo de iones calcio hacia el interior de la fibra es el principal responsable del potencial de acción. Este flujo ocurre de la misma manera autorregenerativa que se produce en los canales de sodio de las fibras nerviosas y de las fibras musculares esqueléticas. Sin embargo, los canales de calcio se abren muchas veces más lentos que los canales de sodio, y también permanecen abiertos mucho más tiempo. Estas características explican en gran medida los prolongados potenciales de acción en meseta de algunas fibras musculares lisas. Excitación del músculo liso visceral por distensión muscular Cuando el músculo liso visceral (unitario) es distendido lo suficiente, habitualmente se generan potenciales de acción espontáneos, que se deben a una combinación de: 1) los potenciales de onda lenta normales, y 2) la disminución de la negatividad global del potencial de membrana que produce la distensión. Las fibras musculares lisas del músculo liso multiunitario (como el músculo del iris del ojo o el músculo erector de cada uno de los cabellos) normalmente se contraen sobre todo en respuesta a estímulos nerviosos. Las terminaciones nerviosas secretan acetilcolina en el caso de algunos músculos lisos multiunitarios y noradrenalina en el caso de otros.
Efectos de las hormonas sobre la contracción del músculo liso Muchas de las hormonas circulantes en la sangre afectan en cierto grado a la contracción del músculo liso, y algunas tienen efectos profundos. Entre las más importantes se encuentran la noradrenalina, la adrenalina, la angiotensina II, la endotelina, la vasopresina, la oxitocina, la serotonina y la histamina. Una hormona produce contracción del músculo liso cuando la membrana de la célula muscular contiene receptores excitadores activados por hormonas para esa hormona.
Aproximadamente el 40% del cuerpo es músculo esquelético, y tal vez otro 10% es músculo liso y cardíaco.
ANATOMIA FISIOLOGICA DEL MUSCULO ESQUELETICO. «Fibras del músculo esquelético«
En la mayoría de los músculos esqueléticos, las fibras se extienden a lo largo de toda la longitud del músculo. Todas las fibras, excepto alrededor de un 2%, habitualmente están inervadas por una sola terminación nerviosa localizada cerca del punto medio de la fibra.
El sarcolema es una fina membrana que envuelve a una fibra musculoesquelética. El sarcolema está formado por una membrana celular verdadera, denominada membrana plasmática, y una cubierta externa formada por una capa delgada de material polisacárido que contiene numerosas fibrillas delgadas de colágeno. Las miofibrillas están formadas por filamentos de actina y miosina Cada fibra muscular contiene varios cientos a varios miles de miofibrillas, los filamentos gruesos de los diagramas son miosina y los filamentos delgados son actina. La relación de yuxtaposición entre los filamentos de miosina y de actina se mantiene por medio de un gran número de moléculas filamentosas de una proteína denominada titina, esta como es muy moleculosa es muy elástica a la vez. Estas moléculas elásticas de titina actúan como armazón que mantiene en su posición los filamentos de miosina y de actina, de modo que funcione la maquinaria contráctil del sarcómero. Un extremo de la molécula de titina es elástico y está unido al disco Z; para actuar a modo de muelle y con una longitud que cambia según el sarcómero se contrae y se relaja. La otra parte de la molécula de titina la une al grueso filamento de miosina. La molécula de titina también parece actuar como molde para la formación inicial de porciones de los filamentos contráctiles del sarcómero, especialmente los filamentos de miosina.
Las muchas miofibrillas de cada fibra muscular están yuxtapuestas suspendidas en la fibra muscular. Los espacios entre las miofibrillas están llenos de un líquido intracelular denominado sarcoplasma, que contiene grandes cantidades de potasio, magnesio y fosfato, además de múltiples enzimas proteicas. También hay muchas mitocondrias que están dispuestas paralelas a las miofibrillas. En el sarcoplasma que rodea a las miofibrillas de todas las fibras musculares también hay un extenso retículo denominado retículo sarcoplasmático.
«Mecanismo general de la contracción muscular« El inicio y la ejecución de la contracción muscular se producen en las siguientes etapas secuenciales: 1. Un potencial de acción viaja a lo largo de una fibra motora hasta sus terminales sobre las fibras musculares. 2. En cada terminal, el nervio secreta una pequeña cantidad de la sustancia neurotransmisora acetilcolina. 3. La acetilcolina actúa en una zona local de la membrana de la fibra muscular para abrir múltiples canales de cationes «activados por acetilcolina» a través de moléculas proteicas que flotan en la membrana. 4. La apertura de los canales activados por acetilcolina permite que grandes cantidades de iones sodio difundan hacia el interior de la membrana de la fibra muscular. Esta acción provoca una despolarización local que, a su vez, conduce a la apertura de los canales de sodio activados por el voltaje, que inicia un potencial de acción en la membrana. 5. El potencial de acción viaja a lo largo de la membrana de la fibra muscular de la misma manera que los potenciales de acción viajan a lo largo de las membranas de las fibras nerviosas. 6. El potencial de acción despolariza la membrana muscular, y buena parte de la electricidad del potencial de acción fluye a través del centro de la fibra muscular, donde hace que el retículo sarcoplásmico libere grandes cantidades de iones calcio que se han almacenado en el interior de este retículo. 7. Los iones calcio inician fuerzas de atracción entre los filamentos de actina y miosina, haciendo que se deslicen unos sobre otros en sentido longitudinal, lo que constituye el proceso contráctil. 8. Después de una fracción de segundo los iones calcio son bombeados de nuevo hacia el retículo sarcoplásmico por una bomba de Ca++ de la membrana y permanecen almacenados en el retículo hasta que llega un nuevo potencial de acción muscular; esta retirada de los iones calcio desde las miofibrillas hace que cese la contracción muscular.
La contracción muscular se produce por un mecanismo de deslizamiento de los filamentos. el mecanismo básico de la contracción muscular, el estado relajado de un sarcómero (arriba) y su estado contraído (abajo). En el estado relajado, los extremos de los filamentos de actina que se extienden entre dos discos Z sucesivos apenas comienzan a superponerse entre sí. Por el contrario, en el estado contraído estos filamentos de actina han sido traccionados hacia dentro entre los filamentos de miosina, de modo que sus extremos se superponen entre sí en su máxima extensión. Además, los discos Z han sido traccionados por los filamentos de actina hasta los extremos de los filamentos de miosina. Así, la contracción muscular se produce por un mecanismo de deslizamiento de los filamentos. La molécula de miosina está formada por seis cadenas polipeptídicas, dos cadenas pesadas, cada una de las cuales tiene un peso molecular de aproximadamente 200.000, y cuatro cadenas ligeras, que tienen un peso molecular de aproximadamente 20.000 cada una. Las dos cadenas pesadas se enrollan entre sí en espiral para formar una hélice doble, que se denomina cola de la molécula de miosina. Un extremo de cada una de estas cadenas se pliega bilateralmente para formar una estructura polipeptídica globular denominada cabeza de la miosina. Así, hay dos cabezas libres en un extremo de la molécula de miosina de doble hélice. Las cuatro cadenas ligeras también forman parte de la cabeza de la miosina, dos en cada cabeza. Estas cadenas ligeras ayudan a controlar la función de la cabeza durante la contracción muscular. Otra característica de la cabeza de la miosina que es esencial para la contracción muscular es que actúa como una enzima adenosina trifosfatasa (ATPasa), el esqueleto del filamento de actina es una molécula de la proteína F-actina bicatenaria. Cada una de las hebras de la doble hélice de F-actina está formada por moléculas de G-actina polimerizadas, cada una de las cuales tiene un peso molecular de aproximadamente 42.000. A cada una de estas moléculas de G-actina se le une una molécula de ADP. Se piensa que estas moléculas de ADP son los puntos activos de los filamentos de actina con los que interactúan los puentes cruzados de los filamentos de miosina para producir la contracción muscular.
Moléculas de tropomiosina El filamento de actina también contiene otra proteína, la tropomiosina. Cada molécula de tropomiosina tiene un peso molecular de 70.000 y una longitud de 40 nm. Estas moléculas están enrolladas en espiral alrededor de los lados de la hélice de F-actina. En estado de reposo las moléculas de tropomiosina recubren los puntos activos de las hebras de actina, de modo que no se puede producir atracción entre los filamentos de actina y de miosina para producir la contracción. Troponina y su función en la contracción muscular Unidas de manera intermitente a lo largo de los lados de las moléculas de tropomiosina hay otras moléculas proteicas denominadas troponina. Estas moléculas proteicas son en realidad complejos de tres subunidades proteicas unidas entre sí de manera laxa, cada una de las cuales tiene una función específica en el control de la contracción muscular. Un filamento de actina puro sin la presencia del complejo troponina-tropomiosina (pero en presencia de iones magnesio y ATP) se une instantánea e intensamente a las cabezas de las moléculas de miosina. Después, si se añade el complejo troponina-tropomiosina al filamento de actina, no se produce la unión entre la miosina y la actina.
ATP como fuente de energía para la contracción: fenómenos químicos en el movimiento de las cabezas de miosina Cuando se contrae el músculo, se realiza un trabajo y es necesaria energía. Durante el proceso de contracción se escinden grandes cantidades de ATP para formar ADP; cuanto mayor sea la magnitud del trabajo que realiza el músculo, mayor será la cantidad de ATP que se escinde, lo que se denomina efecto Fenn. Se piensa que este efecto se produce por medio de la siguiente secuencia de acontecimientos: 1. Antes de que comience la contracción, las cabezas de los puentes cruzados se unen al ATP. La actividad ATPasa de la cabeza de miosina escinde inmediatamente el ATP, aunque deja los productos de la escisión, el ADP y el ion fosfato, unidos a la cabeza. En este estado la conformación de la cabeza es tal que se extiende perpendicularmente hacia el filamento de actina, pero todavía no está unida a ella. 2. Cuando el complejo troponina-tropomiosina se une a los iones calcio quedan al descubierto los puntos activos del filamento de actina, y entonces las cabezas de miosina se unen a estos sitios, como se muestra en la figura 6-8. 3. El enlace entre la cabeza del puente cruzado y el punto activo del filamento de actina produce un cambio conformacional de la cabeza, lo que hace que la cabeza se desplace hacia el brazo del puente cruzado, lo que proporciona el golpe activo para tirar del filamento de actina. La energía que activa el golpe activo es la energía que ya se ha almacenado, como un muelle «comprimido» por el cambio conformacional que se había producido previamente en la cabeza cuando se escindió la molécula de ATP. 4. Una vez que se desplaza la cabeza del puente cruzado, se facilita la liberación del ADP y el ion fosfato que previamente estaban unidos a la cabeza. En el punto de liberación del ADP se une una nueva molécula de ATP, lo cual hace que la cabeza se separe de la actina. 5. Después de que la cabeza se haya separado de la actina, se escinde la nueva molécula de ATP para comenzar el ciclo siguiente, dando lugar a un nuevo golpe activo. Es decir, la energía «comprime» la cabeza una vez más a su situación perpendicular, dispuesta para comenzar el nuevo ciclo de golpe activo. 6. Cuando la cabeza comprimida (con su energía almacenada procedente del ATP escindido) se une a un nuevo punto activo del filamento de actina, se estira y una vez más proporciona un nuevo golpe activo.
Un músculo esquelético se contrae rápidamente cuando lo hace frente a una carga nula, hasta un estado de contracción completa en aproximadamente 0,1 s para un músculo medio. La disminución de la velocidad de contracción al aumentar la carga está producida por el hecho de que una carga sobre un músculo en contracción es una fuerza inversa que se opone a la fuerza contráctil que produce la contracción muscular.
ENERGETICA DE LA CONTRACCION MUSCULAR. Cuando un músculo se contrae contra una carga realiza un trabajo. Esto significa que se transfiere energía desde el músculo hasta la carga externa para levantar un objeto hasta una mayor altura o para superar la resistencia al movimiento. La mayor parte de la energía necesaria para la contracción muscular se utiliza para activar el mecanismo de cremallera mediante el cual los puentes cruzados tiran de los filamentos de actina, aunque son necesarias cantidades pequeñas para: 1) bombear iones calcio desde el sarcoplasma hacia el interior del retículo sarcoplásmico después de que haya finalizado la contracción 2) para bombear iones sodio y potasio a través de la membrana de la fibra muscular para mantener un entorno iónico adecuado para la propagación de los potenciales de acción de la fibra muscular. La contracción de ATP en la fibra muscular, de aproximadamente 4 milimolar, es suficiente para mantener la contracción completa durante solo 1 a 2 s como máximo. La primera fuente de energía que se utiliza para reconstituir el ATP es la sustancia fosfocreatina, que contiene un enlace fosfato de alta energía similar a los enlaces del ATP. La segunda fuente importante de energía, que se utiliza para reconstituir tanto el ATP como la fosfocreatina, es la «glucólisis» del glucógeno que se ha almacenado previamente en las células musculares. La escisión enzimática rápida del glucógeno en ácido pirúvico y ácido láctico libera energía que se utiliza para convertir el ADP en ATP; después se puede utilizar directamente el ATP para aportar energía a la contracción muscular adicional y también para reconstituir los almacenes de fosfocreatina.
Eficiencia de la contracción muscular La eficiencia de una máquina o de un motor se calcula como el porcentaje del aporte de energía que se convierte en trabajo en lugar de en calor. El porcentaje de aporte energético al músculo (la energía química de los nutrientes) que se puede convertir en trabajo, incluso en las mejores condiciones, es menor del 25%, y el resto se convierte en calor. La razón de esta baja eficiencia es que aproximadamente la mitad de la energía de los nutrientes se pierde durante la formación del ATP, y que incluso en este caso solo el 40-45% de la energía del propio ATP se puede convertir posteriormente en trabajo.
CARACTERISTICAS DE LAS CONTRACCIONES MUSCULARES. Esto se puede conseguir con la excitación eléctrica instantánea del nervio que inerva un músculo o haciendo pasar un estímulo eléctrico breve a través del propio músculo, dando lugar a una única contracción súbita que dura una fracción de segundo. Se dice que la contracción muscular es isométrica cuando el músculo no se acorta durante la contracción e isotónica cuando se acorta, pero la tensión del músculo permanece constante durante toda la contracción. En el sistema isométrico, el músculo se contrae contra un transductor de fuerza sin disminuir la longitud del músculo. En el sistema isotónico el músculo se acorta contra una carga fija, que se ilustra en la parte superior de la figura, que muestra un músculo que eleva un peso. Las características de la contracción isotónica dependen de la carga contra la que se contrae el músculo, así como de la inercia de la carga.
Fibras lentas (tipo I, músculo rojo) Las características de las fibras lentas son las siguientes: 1. Las fibras son más pequeñas que las fibras rápidas. 2. Las fibras lentas están también inervadas por fibras nerviosas más pequeñas. 3. En comparación con las fibras rápidas, las fibras lentas tienen un sistema de vascularización más extenso y más capilares para aportar cantidades adicionales de oxígeno. 4. Las fibras lentas tienen números muy elevados de mitocondrias, también para mantener niveles elevados de metabolismo oxidativo. 5. Las fibras lentas contienen grandes cantidades de mioglobina, una proteína que contiene hierro y que es similar a la hemoglobina de los eritrocitos. La mioglobina se combina con el oxígeno y lo almacena hasta que sea necesario, lo cual acelera también notablemente el transporte de oxígeno hacia las mitocondrias. La mioglobina da al músculo lento un aspecto rojizo y el nombre de músculo rojo. Fibras rápidas (tipo II, músculo blanco) Las características de las fibras rápidas son: 1. Las fibras rápidas son grandes para obtener una gran fuerza de contracción. 2. Existe un retículo sarcoplásmico extenso para una liberación rápida de iones calcio al objeto de iniciar la contracción. 3. Están presentes grandes cantidades de enzimas glucolíticas para la liberación rápida de energía por el proceso glucolítico. 4. Las fibras rápidas tienen una vascularización menos extensa que las lentas, porque el metabolismo oxidativo tiene una importancia secundaria. 5. Las fibras rápidas tienen menos mitocondrias que las lentas, también porque el metabolismo oxidativo es secundario. Un déficit de mioglobina roja en el músculo rápido le da el nombre de músculo blanco.
Unidad motora: todas las fibras musculares inervadas por una única fibra nerviosa Todas las motoneuronas que salen de la médula espinal inervan múltiples fibras nerviosas y el número de fibras inervadas depende del tipo de músculo. Todas las fibras musculares que son inervadas por una única fibra nerviosa se denominan unidad motora .
Sumación significa la adición de los espasmos individuales para aumentar la intensidad de la contracción muscular global. La sumación se produce de dos maneras: 1) aumentando el número de unidades motoras que se contraen de manera simultánea, lo que se denomina sumación de fibras múltiples 2) aumentando la frecuencia de la contracción, lo que se denomina sumación de frecuencia y que puede producir tetanización. La máxima fuerza de contracción tetánica de un músculo que funciona a una longitud muscular normal es en promedio de entre 3 y 4 kg por centímetro cuadrado de músculo. Como un músculo cuádriceps puede tener hasta 100 cm2 de vientre muscular, se pueden aplicar hasta 360 kg de tensión al tendón rotuliano. Cuando un músculo comienza a contraerse después de un período de reposo prolongado, su fuerza de contracción inicial puede ser tan pequeña como la mitad de su fuerza entre 10 y 50 contracciones musculares después. Es decir, la fuerza de la contracción aumenta hasta una meseta, un fenómeno que se denomina efecto de la escalera o Treppe. Fatiga muscular La contracción prolongada e intensa de un músculo da lugar al conocido estado de fatiga muscular. Algunos estudios llevados a cabo en atletas han mostrado que la fatiga muscular aumenta en una proporción casi directa a la velocidad de depleción del glucógeno del músculo. Por tanto, la fatiga se debe principalmente a la incapacidad de los procesos contráctiles y metabólicos de las fibras musculares de continuar generando el mismo trabajo.
Hipertrofia y atrofia muscular El aumento de la masa total de un músculo se denomina hipertrofia muscular. Cuando la masa total disminuye, el proceso recibe el nombre de atrofia muscular. Prácticamente toda la hipertrofia muscular se debe a un aumento del número de filamentos de actina y miosina en cada fibra muscular, dando lugar a aumento de tamaño de las fibras musculares individuales; esta situación se denomina hipertrofia de las fibras. La hipertrofia aparece en un grado mucho mayor cuando el músculo está sometido a carga durante el proceso contráctil. Solo son necesarias unas pocas contracciones intensas cada día para producir una hipertrofia significativa en un plazo de 6 a 10 semanas. La ruta que parece importar en buena parte para la degradación proteica en un músculo que experimenta atrofia es la ruta de ubicuitina-proteasoma dependiente del ATP. En la fase final de la atrofia por denervación, la mayor parte de las fibras musculares es destruida y sustituida por tejido fibroso y adiposo. Las fibras que permanecen están formadas por una membrana celular larga con los núcleos de las células musculares alineados, pero con propiedades contráctiles escasas o nulas y con una capacidad escasa o nula de regeneración de las miofibrillas si vuelve a crecer un nervio.
En la fase final de la atrofia por denervación, la mayor parte de las fibras musculares es destruida y sustituida por tejido fibroso y adiposo. Las fibras que permanecen están formadas por una membrana celular larga con los núcleos de las células musculares alineados, pero con propiedades contráctiles escasas o nulas y con una capacidad escasa o nula de regeneración de las miofibrillas si vuelve a crecer un nervio.